






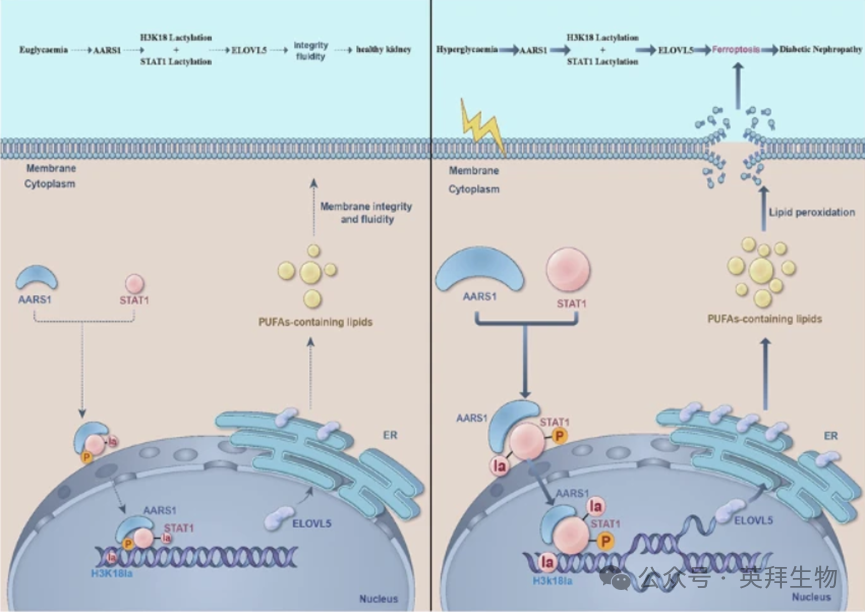
AARS1 介导的 H3K18 和 STAT1 的乳酸化作用可促进糖尿病肾病中的铁死亡
糖尿病肾病(DN)是全球范围内导致终末期肾病的主要原因。近期研究表明,由乳酸介导的组蛋白乳酸化(作为一种新型的表观遗传修饰)参与了糖尿病相关并发症的发生和发展过程。然而,关于乳酰转移酶在糖尿病肾病中的作用知之甚少。丙氨酰-核糖胺合成酶 1(AARS1)被鉴定为一种新型的乳酰转移酶,能够调节组蛋白 H3-赖氨酸-18 乳酸化(H3K18la)。在本研究中,作者确定了 AARS1 介导的 H3K18la 是否参与糖尿病肾病的发病机制。更重要的是,作者探索了其中可能涉及的机制。本研究中使用了由野生型和丙氨酰-核糖胺合成酶(AARS1)杂合子(AARS1+/-)小鼠组成的糖尿病肾病模型。通过转录组学和脂质组学分析,并结合多种分子生物学方法,作者试图阐明 AARS1 如何通过调节糖尿病肾病中的铁死亡来调节肾脏功能和肾细胞死亡的潜在机制。作者的研究结果表明,AARS1 和 H3K18la 表达的增加通过调节糖尿病肾病模型中的铁死亡参与了肾脏功能障碍和肾细胞死亡。此外,AARS1 通过增加脂肪酸延长酶-5(ELOVL5)的转录来引发脂质过氧化,最终导致铁死亡的诱导。此外,AARS1 与信号转导及转录激活因子 1(STAT1)相互作用,共同调节 ELOVL5 的转录。另外,使用 STAT1 特异性抑制剂氟达拉滨可延缓 DN 的进展。此外,作者观察到 AARS1 调节 STAT1 和 H3K18 的乳酸化以调节 ELOVL5 的转录,从而触发铁死亡。通过β-丙氨酸抑制 AARS1 诱导的乳酸化,可减轻 DN 模型小鼠和高血糖细胞中的铁死亡。本研究表明,AARS1 诱导 H3K18 和 STAT1 的乳酸化以调节 ELOVL5 的转录,从而在糖尿病肾病模型中触发铁死亡。该文章于2026年3月发表在《Cell death and differentiation》,影响因子:15.4。
技术路线:
主要研究结果:
1. 在糖尿病肾病(DN)患者及模型中,AARS1 和 H3K18la 的表达水平均有所升高
本研究纳入的 DN 患者的特点如补充表 1 所示。这些 DN 患者根据估算的肾小球滤过率(eGFR)被分为 2 期、3 期或 4 期。DN 患者的样本经苏木精-伊红(HE)染色后,其结果如图 1A 所示。马松三色染色显示随着 DN 病程的进展,胶原沉积和间质纤维化程度加重(图 1A)。同样地,TUNEL 检测结果表明,随着 DN 病程的进展,DN 患者肾脏中的细胞死亡增加(图 1A)。此外,在肾小管和肾小球中均观察到细胞死亡(图 1A)。先前的研究表明,H3K18la 可能参与糖尿病相关并发症的发生和发展。此外,AARS1 被证实为一种新型的乳酰转移酶,能够调节 H3K18la。因此,作者检测了 DN 患者肾脏中 AARS1、乳酸化和 H3K18la 的水平。作者的数据表明,在糖尿病肾病患者的肾脏中,随着糖尿病肾病病情的发展,AARS1 的水平、乳酸化程度以及 H3K18la 的含量均有所上升(图 1A)。
本研究中所用小鼠的血清生化指标见附表 2。HE 和马松三色染色显示,糖尿病肾病小鼠的肾脏出现了显著变化(图 1B)。TUNEL 染色表明,糖尿病肾病小鼠的肾脏细胞死亡有所增加(图 1B)。此外,糖尿病肾病小鼠肾脏中 AARS1、乳化作用和 H3K18la 的蛋白水平均有所升高(图 1B、C)。同时,糖尿病肾病小鼠肾脏中 AARS1 的 mRNA 水平也有所增加(图 1D)。
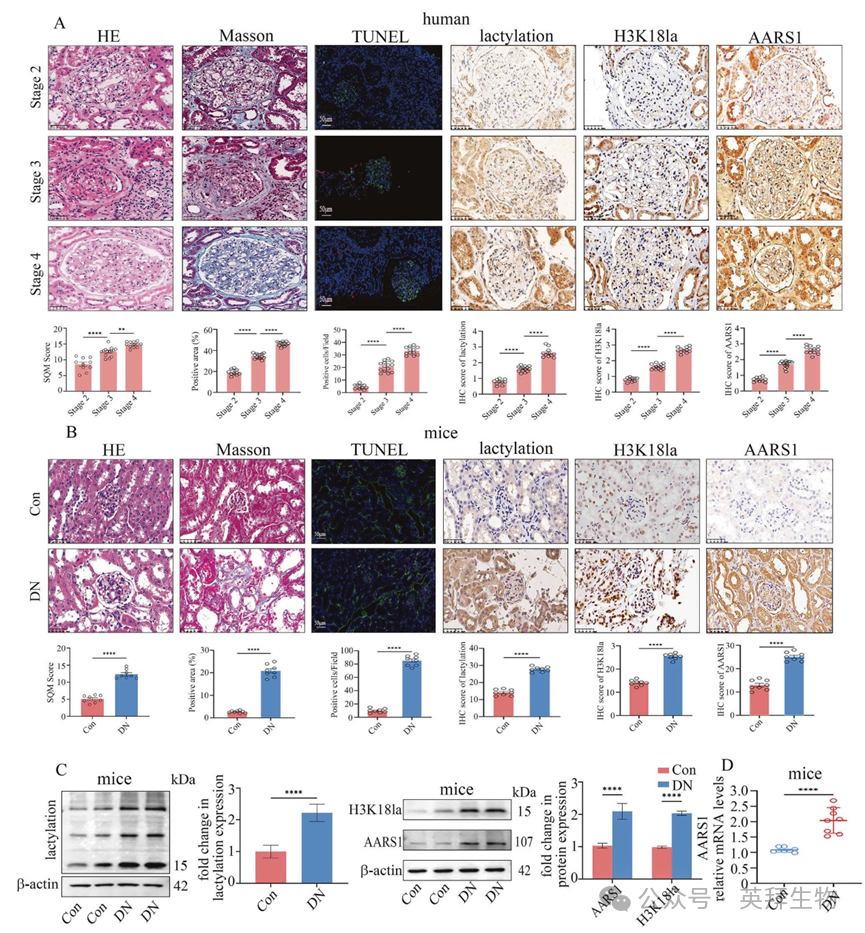
图 1 糖尿病肾病(DN)患者及小鼠体内 AARS1 和 H3K18la 的表达水平均有所升高
此外,高糖处理使人类肾小球内皮细胞(HGECs)和人类肾小管上皮细胞(HK-2)中的 AARS1、乳化作用以及 H3K18la 水平升高(补充图 1A、B)。而且,高血糖细胞中的细胞死亡情况也有所增加(补充图 1C)。此外,作为渗透控制剂的甘露醇处理对高血糖细胞中的 AARS1、乳化作用或 H3K18la 水平以及细胞死亡没有影响(补充图 1A - C)。这些数据表明,在糖尿病肾病患者和模型中,AARS1、乳化作用和 H3K18la 的水平有所升高。
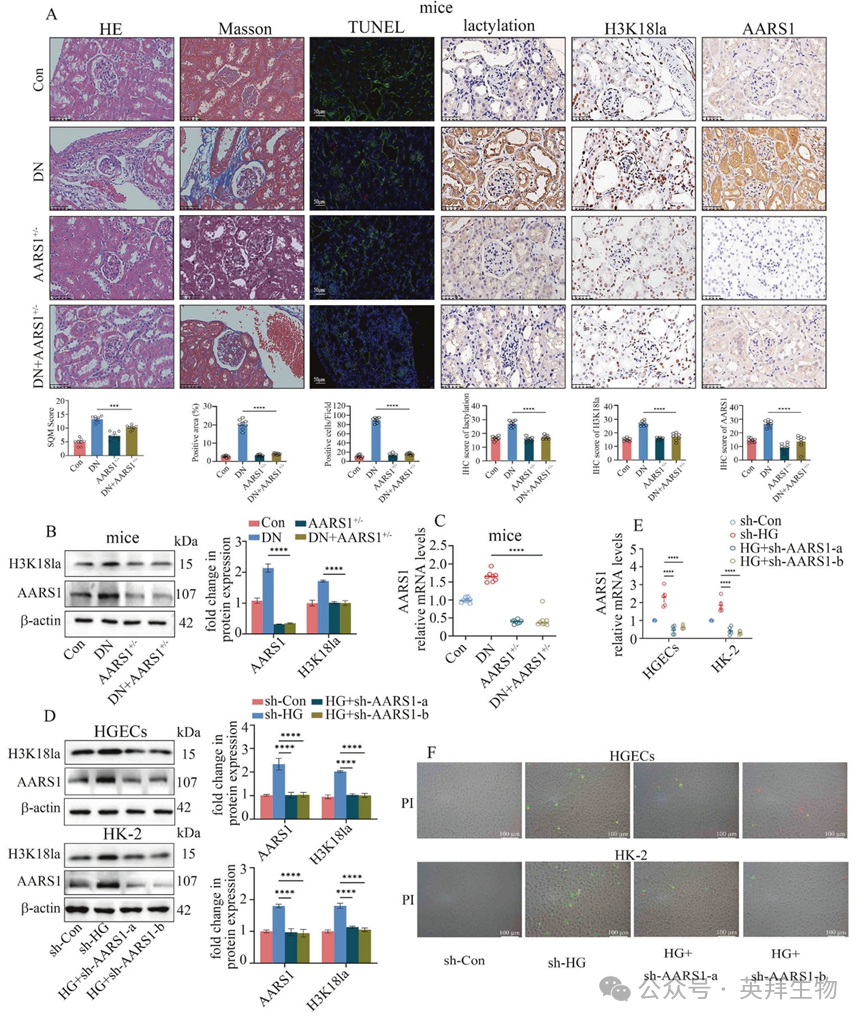
图 2 AARS1的上调参与了糖尿病肾病(DN)的发展过程
2. AARS1 的上调参与了糖尿病肾病(DN)的发展
作者利用 AARS1 杂合型(AARS1+/–)小鼠建立了糖尿病肾病模型(AARS1+/-DN 小鼠),并进一步探究了 AARS1 在糖尿病肾病中的作用(图 2A–C)。与糖尿病小鼠相比,AARS1+/-DN 小鼠的肾脏组织结构损伤更轻;纤维化减轻;肾细胞死亡减少;AARS1、乳酸化和 H3K18la 水平降低(图 2A‒C);并且肾功能得到改善(补充表 2)。与这些发现一致的是,沉默 AARS1 降低了高血糖细胞中 AARS1 和 H3K18la 的表达(图 2D、E)。此外,沉默 AARS1 逆转了高糖介导的 HGECs 和 HK-2 细胞的死亡(图 2F)。此外,Gln-AMS,一种 AARS1 抑制剂,以浓度和时间依赖的方式抑制 AARS1 和 H3K18la 的表达(补充图 1D、E)。此外,Gln-AMS 治疗降低了高血糖细胞的死亡(补充图 1F)。这些数据表明,AARS1 诱导的 H3K18la 参与了糖尿病肾病的发展。
3. AARS1 的上调参与了糖尿病肾病(DN)的发病机制,其作用机制是通过触发铁死亡来实现的
作者对小鼠肾脏进行了 RNA-测序分析,以进一步探究 AARS1 诱导的 H3K18la 参与 DN 发病机制的具体方式。作者的研究结果表明,在患有 DN 的小鼠肾脏中,与不饱和脂肪酸的生物合成、脂肪酸延长、脂肪酸代谢以及代谢途径相关的 KEGG 通路得到了丰富(图 3A,补充表 3)。此前的研究表明,PUFA 生物合成途径在铁死亡中起着关键作用。同样,在 DN 患者的肾脏中,4-羟基-2-壬烯醛(4-HNE)、丙二醛(MDA)和长链酰基辅酶 A 合成酶家族成员 4(ACSL4)的水平逐渐升高,而谷胱甘肽过氧化物酶 4(GPX4)的水平则逐渐降低,随着 DN 阶段的进展(图 3B),这表明铁死亡参与了 DN 的发展。作者进一步验证了铁死亡是否参与了 DN 的进展,通过用铁死亡抑制剂铁丝汀-1(Fer-1)处理患有 DN 的小鼠和高糖暴露的细胞来实现。Fer-1 能减轻肾脏组织损伤,抑制 ACSL4 的表达,并提高 GPX4 的水平(补充图 2A、B)。此外,通过透射电子显微镜(TEM)观察发现,DN 小鼠的铁死亡特征,如具有明显减少的线粒体嵴和较小体积的线粒体的密集线粒体结构,在接受 Fer-1 处理后有所减轻(补充图 2A)。此外,Fer-1 还减少了 DN 小鼠肾脏中的 MDA 和脂质过氧化物(LPO)沉积,降低了 Fe2+含量,并改善了肾脏功能障碍(补充图 2C - E;补充表 2)。在高血糖细胞中,Fer-1 以时间和浓度依赖的方式减少了高血糖细胞的死亡(补充图 3A)。此外,Fer-1 在高血糖细胞中降低了 ACSL4 的表达,并提高了 GPX4 的水平(补充图 3B)。C11-BODIPY 581/591(一种脂质过氧化探针)荧光染色显示,Fer-1 抑制了高糖介导的脂质过氧化(补充图 3C)。JC-1 是一种线粒体膜电位(MMP)探针,荧光染色结果显示,Fer-1 处理能够改善高糖诱导的 HGECs 和 HK-2 细胞中 MMP 的破坏情况(补充图 3D)。Fer-1 处理降低了高糖诱导的丙二醛(MDA)水平(补充图 3E)。针对 FeRhoNox-1(一种特定的 Fe2+探针)的荧光染色显示,Fer-1 处理降低了高血糖细胞中的 Fe2+含量(补充图 3F)。针对线粒体特异性超氧化物指示剂 MitoSOX Red 的荧光染色表明,Fer-1 处理减少了线粒体超氧化物的积累(补充图 3G)。
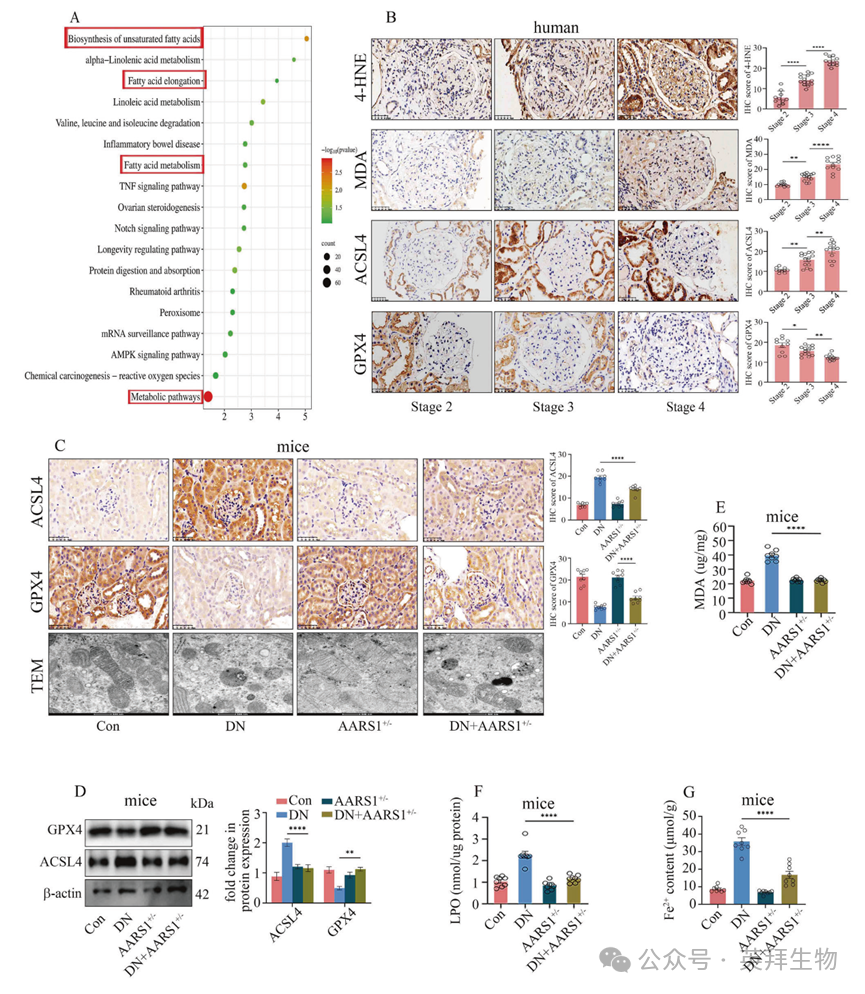
图 3 AARS1的上调参与了糖尿病肾病(DN)的发病机制,其作用方式是通过引发铁死亡来实现的
作者在糖尿病肾病模型中检测到了与铁死亡相关的指标,以进一步证实 AARS1 诱导的 H3K18la 对糖尿病肾病中的铁死亡具有调节作用。作者的数据表明,ACSL4 的表达降低,GPX4 的水平升高,在 TEM 图像中铁死亡得到改善,MDA 和 LPO 的含量降低,肾脏中的 Fe2+含量在 AARS1+/– 糖尿病肾病小鼠中低于糖尿病小鼠(图 3C–G)。同样,在高血糖细胞(HGECs)和 HK-2 细胞中,铁死亡标志物的水平升高,而通过 AARS1 基因沉默可使其逆转(补充图 4)。一致地,Gln-AMS 处理抑制了高血糖细胞中的铁死亡(补充图 5)。这些数据表明,AARS1 诱导的 H3K18la 表达通过触发铁死亡参与了糖尿病肾病的发生。
4. AARS1 诱导的 H3K18la 参与了糖尿病肾病(DN)模型中的铁死亡过程,其机制是通过调节 ELOVL5 的转录实现的
接下来,作者探究了 AARS1 诱导的 H3K18la 如何在患有 DN 的个体中参与铁死亡的具体机制。RNA-seq 分析显示,ELOVL5、转-2,3-烯酰辅酶 A 还原酶(TECR)和 3-羟基酰辅酶 A 脱水酶 1(HACD1)参与了多个途径,包括不饱和脂肪酸的生物合成、脂肪酸代谢、脂肪酸延长和代谢途径(补充图 6A、补充表 3)。在这三个基因中,只有 ELOVL5 存在于 H3K18la ChIP-seq 结果中(补充表 4)。ChIP 实验证实,AARS1 和 H3K18la仅通过引物 3(P3)在 ELOVL5 的启动子区域富集,并且再 ChIP 实验表明,AARS1 和 H3K18la 占据了 ELOVL5 启动子的同一区域(补充图 6B)。随着 DN 阶段的进展,DN 患者肾脏中的 ELOVL5 表达水平逐渐升高(补充图 6C)。一致地,DN 模型小鼠(补充图 6D-F)和高血糖细胞(补充图 6G、H)中 ELOVL5 的表达也上调。ELOVL5 表达的抑制减弱了高血糖状态下 HGECs 和 HK-2 细胞中的铁死亡现象(见补充图 7)。对高糖处理的 ELOVL5 基因沉默细胞进行脂质代谢分析后发现,其产生的 PUFAs(包括花生四烯酸 AA)有所减少(补充图 8A、补充表 5)。此外,免疫荧光数据表明,ELOVL5 主要位于内质网中(补充图 8B)。而且,用 AA 处理 ELOVL5 基因沉默的细胞逆转了 si-ELOVL5 对高糖介导的铁死亡的保护作用(补充图 8C - H)。此外,ELOVL5 基因沉默还减弱了由 FINO2(一种铁死亡诱导剂)介导的铁死亡(补充图 9)。这些数据表明,在患有糖尿病的个体中,ELOVL5 通过调节内质网中的 PUFAs 合成来触发铁死亡。
接下来,作者通过调控 ELOVL5 的转录来确定 AARS1 是否能介导糖尿病肾病(DN)患者的铁死亡。作者的数据表明,AARS1 的过表达增加了 ELOVL5 的表达(图 4A、B),并促进了细胞中的铁死亡(图 4)。此外,AARS1 过表达对铁死亡的影响在 ELOVL5 基因沉默的情况下被消除(图 4)。此外,AARS1 表达的抑制降低了糖尿病模型小鼠肾脏中的 ELOVL5 水平(补充图 10A - C)以及高血糖细胞中的 ELOVL5 水平(补充图 10D - G)。这些数据表明,AARS1 通过增加 ELOVL5 的转录来促进铁死亡,在糖尿病模型中发挥着作用。
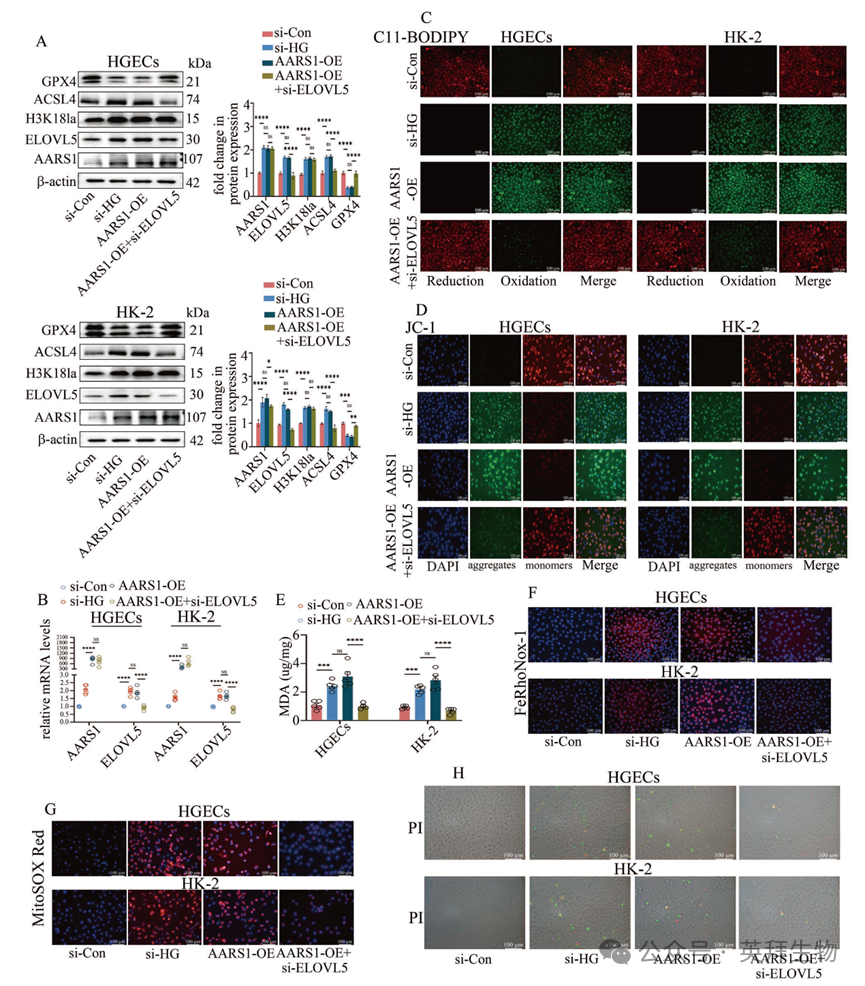
图 4 AARS1调节 ELOVL5 的转录过程,从而在高血糖细胞中介导铁死亡。
5. AARS1 在体内和体外均与信号转导及转录激活因子 1(STAT1)直接相互作用
基因启动子区域的组蛋白修饰通常与转录因子结合相吻合,从而导致下游基因的转录。在本研究中,作者进行了质谱分析以确定与 AARS1 相互作用的转录因子。作者的数据表明,STAT1 可能与 AARS1 相互作用(图 5A,补充表 6)。共免疫沉淀和免疫荧光结果显示,STAT1 在 HGECs 和 HK-2 细胞中与 AARS1 结合(图 5B,C),而 AARS1 与 STAT1 之间的直接结合进一步通过 GST 拉下实验得到证实(图 5D)。此外,作者还评估了 DN 患者和 DN 模型中肾活检样本中的 STAT1 表达水平。作者的数据表明,随着 DN 阶段的进展,STAT1 水平逐渐升高(图 5E)。一致地,DN 模型小鼠肾脏中的 STAT1 表达以及高血糖细胞中的 STAT1 表达均上调(图 5F–H,图 5I,J)。
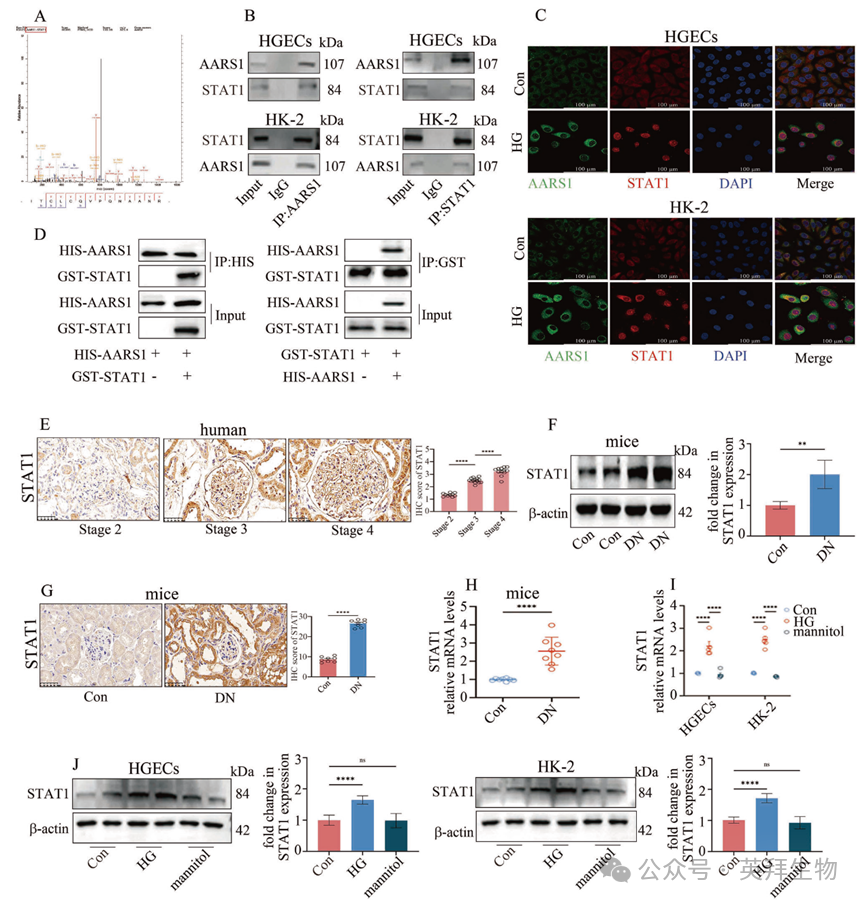
图 5 AARS1与 STAT1 直接相互作用
6. STAT1 通过调节 DN 模型中的 ELOVL5 转录参与铁死亡过程
作者进行了功能缺失和功能增强实验,以确定 STAT1 对 ELOVL5 表达和铁死亡的影响。STAT1 基因沉默降低了 ELOVL5 的水平,并抑制了高血糖 HGECs 和 HK-2 细胞中的铁死亡(图 11 的补充图)。此外,STAT1 过表达上调了 ELOVL5 的表达,并在 HGECs 和 HK-2 细胞中激活了铁死亡,而这些变化通过 ELOVL5 基因沉默而被消除(图 12 的补充图)。本研究中进行了 ChIP 和荧光素酶测定,以确定 STAT1 是否直接调节 ELOVL5 转录。ChIP 测定显示 STAT1 在 ELOVL5 启动子中富集,而再 ChIP 测定显示 STAT1 和 AARS1 占据了 ELOVL5 启动子的同一区域(图 13A 的补充图)。接下来,使用 JASPAR(https://jaspar.genereg.net/)预测了 STAT1 在 ELOVL5 启动子区域的潜在结合位点,结果见补充表 7。位点特异性模式图和位置权重矩阵分别位于上部和下部(图 13B 的补充图)。随后,作者构建了一个双结合位点发生突变的质粒,并发现当这些位点发生突变时,ELOVL5 的转录会被显著抑制(补充图 13C)。因此,作者推测 STAT1 直接与 ELOVL5 的启动子区域结合,并参与了 ELOVL5 转录的调节过程。
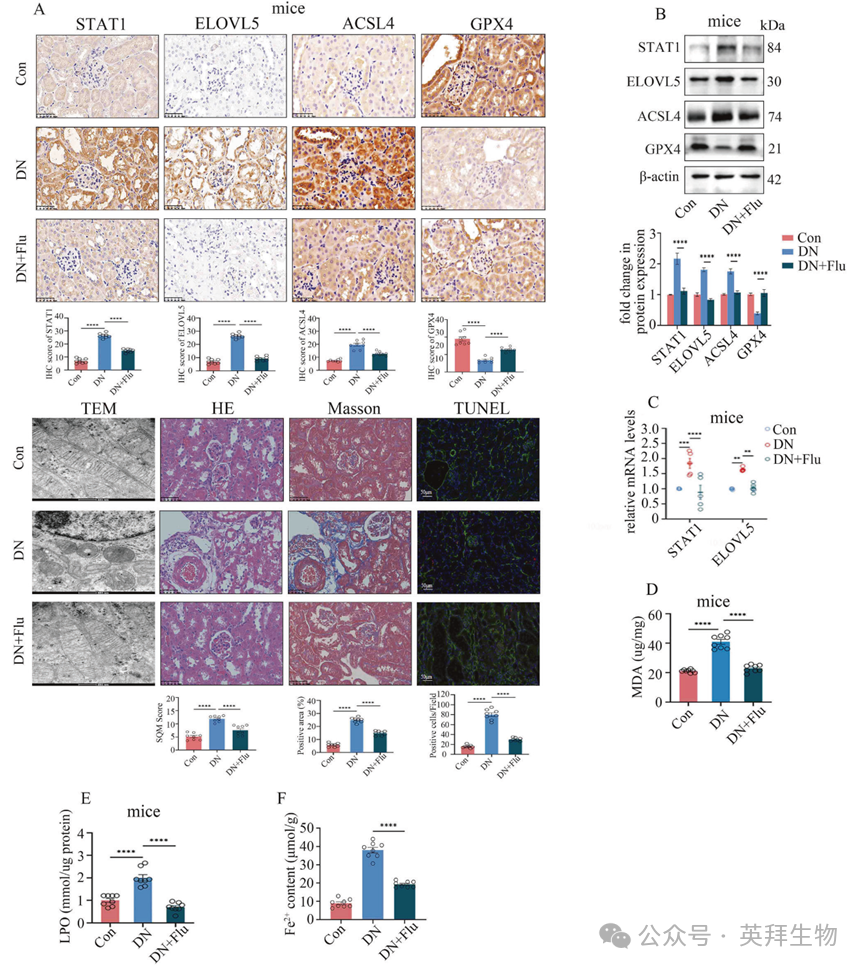
图 6 STAT1抑制剂氟达拉滨(Flu)能够抑制铁死亡,并改善糖尿病肾病(DN)模型小鼠的肾功能障碍
作者进一步探究了 STAT1 是否是抑制 DN 中铁死亡的潜在靶点。在细胞实验中,作者使用了氟达拉滨(Flu),这是一种针对 STAT1 的特异性抑制剂。研究结果表明,氟达拉滨以浓度和时间依赖的方式抑制了 STAT1 的表达(补充图 14A)。此外,氟达拉滨不仅抑制了 STAT1 的表达,还降低了 ELOVL5 的水平(补充图 14B、C),从而在下游抑制了铁死亡(补充图 14B - I)。一致地,在 DN 模型小鼠的腹腔注射中,氟达拉滨有效地下调了 STAT1 和 ELOVL5 的表达(图 6A - C),阻碍了铁死亡(图 6A - F),减轻了肾脏损伤(图 6A),并显著改善了肾脏功能(补充表 2)。这些数据表明,STAT1 抑制剂在预防 DN 方面具有相当大的临床价值。
7. AARS1 调节 STAT1 和 H3K18 的乳酸化水平,从而调控 ELOVL5 的转录并引发铁死亡
接下来,作者探究了 AARS1 是否会影响 STAT1 的转录活性。作者的数据表明,AARS1 的过表达促进了 STAT1 的核转移(图 7A)、增加了 STAT1 的磷酸化(图 7B)以及增强了 STAT1 与 ELOVL5 启动子区域的结合(图 7C)。然而,缺乏乳酰转移酶活性的 AARS15M()并未产生这些效应(图 7A–C)。这些数据表明,AARS1 可能通过调节 STAT1 的乳酸化来增加其转录活性。本研究中采用了体外和体内乳酸化实验来验证作者的假设。作者的体外乳酸化研究表明,AARS1 以浓度依赖的方式利用乳酸诱导 STAT1 乳酸化(图 7D)。AARS1 也被证实能在 HGECs 和 HK-2 细胞中诱导 STAT1 乳酸化(图 7E)。此外,AARS1 过表达增加了 STAT1、H3K18la 和 ELOVL5 的表达(图 7E),并促进了 HGECs 和 HK-2 细胞中的铁死亡(补充图 15)。然而,AARS15M 并未具有这些效应(图 7E;补充图 15)。作者通过改变 STAT1 的乳化位点进一步阐明了 AARS1 介导的 STAT1 乳化在 STAT1 转录功能中的关键作用,正如之前研究中所报道的那样。作者的数据表明,STAT1K193 位点是受 AARS1 调控的最重要的乳化位点(补充图 16A)。此外,STAT1K193 位点的突变逆转了 AARS1 过表达引起的 STAT1 核转位的促进作用、STAT1 磷酸化的增加以及 STAT1 与 ELOVL5 启动子区域结合的增强(补充图 16B–D)。这些发现表明,在高糖环境中,AARS1 在 K193 位点乳化 STAT1,从而增加 STAT1 转录活性。作者的数据表明,AARS1 调节 STAT1 和 H3K18 的乳化以调节 ELOVL5 转录,从而在 DN 中触发铁死亡。
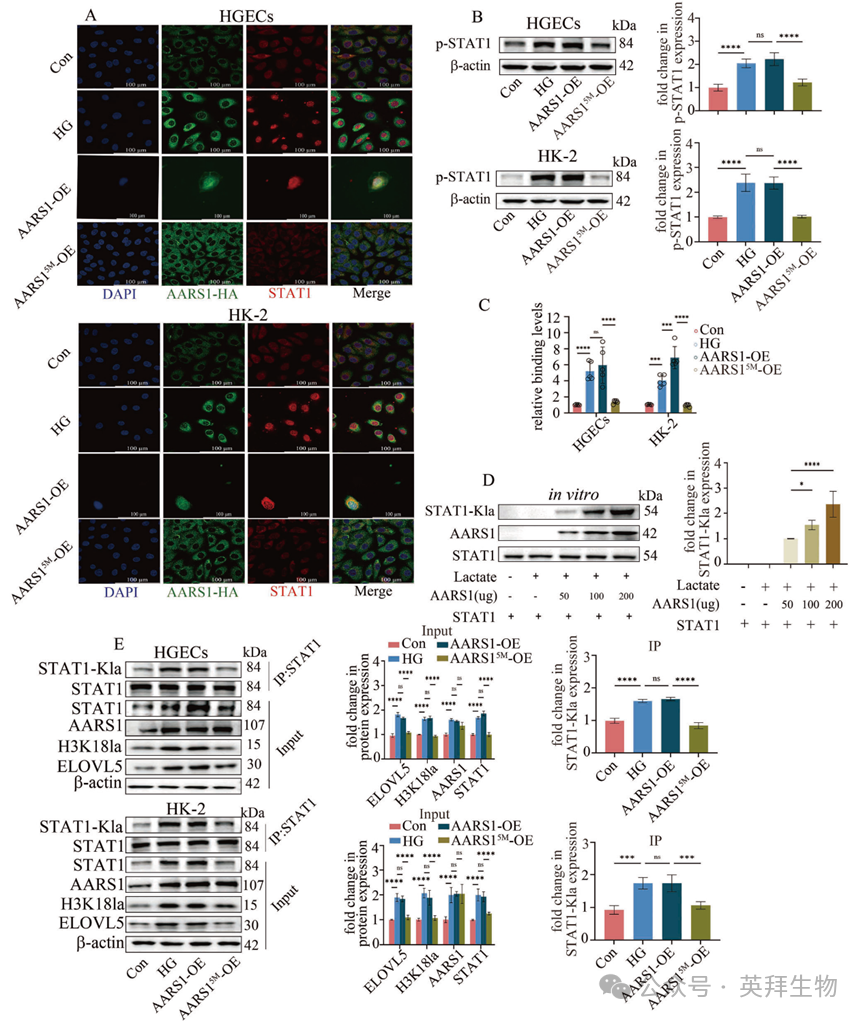
图 7 AARS1调节 STAT1 和 H3K18 的乳酸化过程,从而调控 ELOVL5 的转录并引发铁死亡
8. β-天冬氨酸通过抑制 AARS1 诱导的乳化作用,减轻了糖尿病肾病模型小鼠和高血糖细胞中的铁死亡
鉴于 AARS1 介导的乳化在糖尿病肾病中起着重要作用,抑制 AARS1 介导的乳化可能是治疗糖尿病肾病的一个潜在靶点。此前的一项研究表明,β-天冬氨酸能够通过直接与乳酸竞争与 AARS1 的结合来有效拮抗 AARS1 诱导的乳化作用。
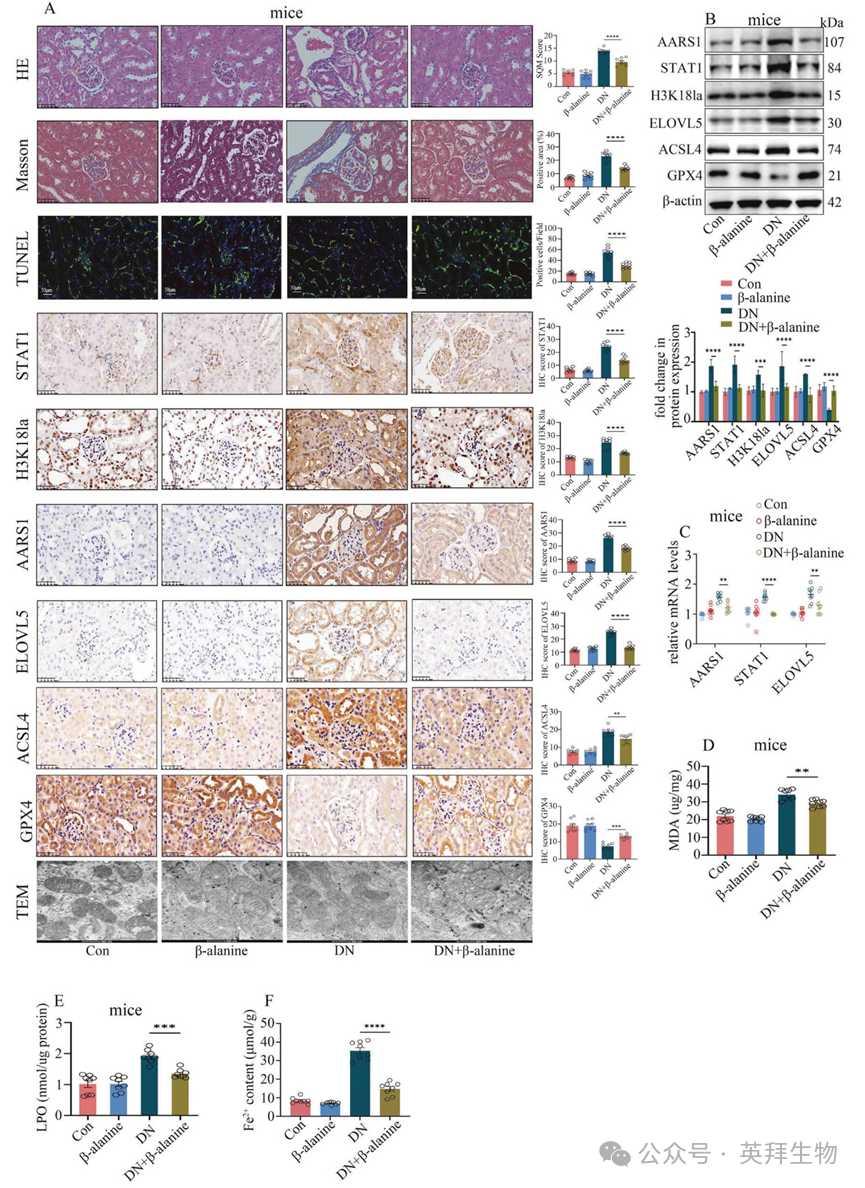
图 8 β-丙氨酸通过抑制 AARS1 诱导的乳化作用,减轻了 DN 模型小鼠体内的铁死亡现象
作者的体外乳化研究证实,β-天冬氨酸能够拮抗 AARS1 诱导的 H3K18 和 STAT1 乳化(补充图 17A、B)。接下来,在本研究中,作者使用β-天冬氨酸对糖尿病模型进行了治疗。作者的数据表明,β-天冬氨酸在高血糖细胞中以浓度和时间依赖的方式抑制高糖诱导的 STAT1 和 H3K18 乳化(补充图 17C)。此外,β-天冬氨酸治疗降低了 AARS1、STAT1、H3K18la 和 ELOVL5 的表达,并抑制了高血糖细胞中的铁死亡(补充图 18)。一致地,β-丙氨酸治疗降低了 AARS1、STAT1、H3K18la 和 ELOVL5 的表达(图 8A–C),抑制了铁死亡(图 8A–F),减轻了肾脏损伤(图 8A),并改善了肾功能障碍(补充表 2),在糖尿病肾病模型小鼠中均表现出了这些效果。这些数据表明,通过β-丙氨酸抑制 AARS1 诱导的乳化作用可能是治疗糖尿病肾病的有效策略。
结论:
ELOVL5 能调节多不饱和脂肪酸的生成,从而引发患有糖尿病肾病(DN)患者的铁死亡。此外,AARS1 起到乳酰转移酶的作用,通过调节 H3K18 和 STAT1K193 的乳酸化来调节 ELOVL5 的转录,从而介导患有糖尿病肾病患者的铁死亡。另外,抑制 AARS1 诱导的乳酸化可能是治疗糖尿病肾病的有效策略。
参考文献:
Hong J, Xu H, Yu L, Yu Z, Chen X, Meng Z, Zhu J, Li J, Zhu M. AARS1-mediated lactylation of H3K18 and STAT1 promotes ferroptosis in diabetic nephropathy. Cell Death Differ. 2026 Mar;33(3):589-604. doi: 10.1038/s41418-025-01587-4.