






lncRNA新发现:PART1与ADAMTS9-AS2拮抗调控前列腺癌AR信号及细胞衰老机制
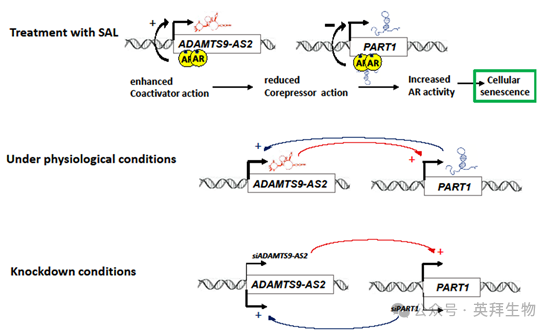
前列腺癌(PCa)最初是一种激素依赖性疾病,其发展和扩散与雄激素受体(AR)信号活性紧密相关。针对 AR 通路的治疗,如使用 AR 拮抗剂和双极雄激素疗法中采用的超生理雄激素水平(SAL),是治疗 PCa 的标准方法。在此,作者通过检测 PCa 标本、观察对 AR 拮抗剂或 SAL 的反应,以及进行功能缺失实验,发现两种长链非编码 RNA(lncRNA)PART1 和 ADAMTS9-AS2 在一定程度上介导了 PCa 细胞中的雄激素信号传导。结果显示PART1 和 ERVH48-1 在 TT 样本中显著高表达,而 ADAMTS9-AS2 在 TT 样本中的表达低于 NTAT 样本。SAL 处理在两种人类 PCa 细胞系和患者肿瘤样本中呈现出相反的调控作用:ADAMTS9-AS2 表达上调,PART1 表达受抑制。此外,研究数据表明,ADAMTS9-AS2 可能作为 AR 信号的共激活因子,而 PART1 则作为共抑制因子。有趣的是,敲低实验显示,ADAMTS9-AS2 和 PART1 均能调节 AR 活性、蛋白水平,以及 SAL 介导的细胞衰老诱导过程。因此,数据表明 ADAMTS9-AS2 和 PART1 在 SAL 条件下对 AR 信号传导起到调控作用。这篇文章于2025年3月发表于《International Journal of Surgery》期刊上,IF:12.5。
研究技术路线:
主要实验结果:
1、患者前列腺癌样本中的差异表达
PART1 在肿瘤组织(TT)与正常对照(NC)之间,以及 TT 与肿瘤旁正常组织(NTAT)之间的表达存在差异(图 1)。ADAMTS9-AS2 在 NTAT 样本中的表达高于 TT 和 NC 样本。C1QTNF3-AMACR 在三组样本中的表达没有差异。C8orf34-AS1 在 NC 样本中的表达高于其他组样本。ERVH48-1在所有三组比较(TT/NC、TT/NTAT 和 NC/NTAT)中的表达均存在差异。最后,KIAA0087 和 ENO1-AS1 在 TT 与 NC 之间,以及 NC 与 NTAT 之间的表达水平不同(图 1)。
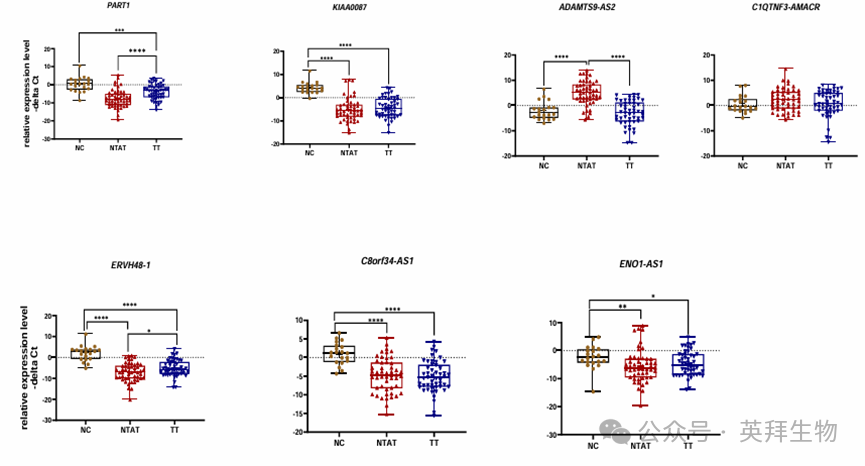
图 1. 比较正常对照组织(NC)、肿瘤旁正常组织(NTAT)和前列腺肿瘤组织(TT)中所选 lncRNA 基因的不同表达模式
PART1 和 ERVH48-1 在 TT 样本中的表达水平高于 NTAT 样本(表达比值分别为13.1 和 3.6;P 值分别 < 0.0001 和 0.04)。值得注意的是,PART1、KIAA0087、ERVH48-1、C8orf34-AS1 和 ENO1-AS1 在 TT 样本中的表达低于 NC 样本(P 值分别为 0.0002、<0.0001、<0.0001、<0.0001 和 0.04)。虽然 PART1、KIAA0087、ERVH48-1、C8orf34-AS1 和 ENO1-AS1 在 NTAT 样本中的表达低于 NC 样本,但 ADAMTS9-AS2 在 NTAT 样本中的表达高于 NC 样本(表达比值(95% 置信区间) = 135(27.3 - 670),P 值 < 0.0001)。详细统计数据见表 S4。这些数据表明,除C1QTNF3-AMACR 外,分析的基因在这些样本中均表现出显著不同的表达水平。基于 NTAT 和 NC 之间三个基因表达的显著差异,数据还表明,NTAT 样本衍生的数据不能总是被视为非肿瘤对照。
在 TT、NTAT 和 NC 样本中,PART1、KIAA0087、ADAMTS9-AS2、C1QTNF3-AMACR、ERVH48-1、C8orf34-AS1 和 ENO1-AS1 的表达水平之间存在显著的成对相关性。最强的相关性出现在 ENO1-ASI和 KIAA0087 之间(相关系数 = 0.95);以及 C8orf34-AS1 与另外两个 lncRNA,即 KIAA0087 和 ERVH48-1 在 TT 样本中的相关性(相关系数 = 0.95)。同样,在这些样本中,ENO1-ASI 的表达水平与 PART1 和 C8orf34-AS1 的表达水平相关,相关系数为 0.94。然后,作者使用层次聚类方法(Ward 法,欧氏距离)根据表达水平将lncRNA 分为四个簇。C1QTNF3-AMACR和 ADAMTS9-AS2 聚为一组;而 ERVH48-1 和 C8orf34-AS1 被识别为聚在另一组。最后,PART1 和 KIAA0087 构成了另一个簇。值得注意的是,大多数患者样本(TT1-TT50 和 NTAT1-NTAT50)中分析的 lncRNA 表达水平降低。
受试者工作特征(ROC)曲线分析显示,KIAA0087在区分 TT 与 NC 样本(AUC = 0.96)以及 NTAT 与 NC 样本(AUC = 0.93)方面表现出色。此外,ERVH48-1 在区分 TT 样本与 NC 样本(AUC = 0.91)以及 NTAT 样本与 NC 样本(AUC = 0.95)方面表现适宜。TT 样本中 C1QTNF3-AMACR 的相对表达水平与体重指数(BMI)相关(P = 0.02)。此外,ADAMTS9-AS2 的表达水平与前列腺体积相关(P = 0.05)。前列腺癌患者的年龄与前列腺体积之间存在显著正相关(x2=6.41),P = 0.01)。正如预期的那样, Gleason 评分与前列腺特异性抗原(PSA)浓度之间也存在显著正相关(=6.28,P = 0.04)。
2、SAL诱导ADAMTS9-AS2上调,而PART1下调
去势抵抗性 C4-2 细胞的实验显示,用 SAL 处理后,ADAMTS9-AS2 显著上调。用第二代 AR 拮抗剂恩杂鲁胺(ENZ)和达罗他胺(ODM)处理对该 lncRNA 的表达没有显著影响(图 2)。另一方面,用 SAL 处理后,PART1 的表达下调,用 AR 拮抗剂 ENZ 或 ODM 处理也会使其下调。通过分析作者之前的 RNA 测序数据,证实了 SAL 对 PART1 的下调作用。
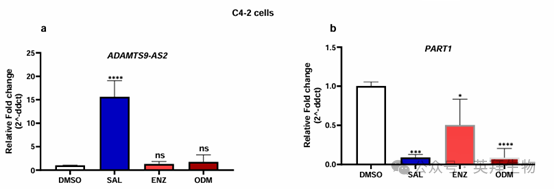
图 2. SAL 处理增强了 lncRNA ADAMTS9-AS2 的表达,而 lncRNA PART1 的表达则被 SAL 和第二代 AR 拮抗剂抑制
这表明这两个 lncRNA 是 AR 信号通路的一部分,也表明AR 拮抗剂并不总是表现出与雄激素相反的调节作用,并且对于 PART1 而言,AR 拮抗剂可能不会使 AR 失活,反而可能赋予 AR 转录活性。
在雄激素敏感的 LNCaP 细胞中进行的类似实验显示,用 SAL 处理可有效诱导ADAMTS9-AS2 的表达。用 Enz 或 ODM 处理也观察到表达上调(图 3)。相反,用 SAL 或 AR 拮抗剂处理后,LNCaP 细胞中 PART1 的表达下调。因此,数据表明 PART1 和 ADAMTS9-AS2 的表达处于 AR 信号通路中。有趣的是,PART1 的表达被 SAL 或 AR 拮抗剂抑制,而 ADAMTS9-AS2 的表达则被 AR 配体(包括 AR 拮抗剂)激活。
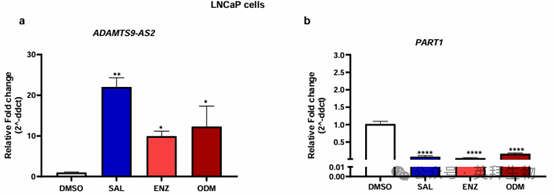
图 3. SAL 和 AR 拮抗剂增强了 lncRNA ADAMTS9-AS2 的表达,而 lncRNA PART1 的表达则被 SAL 和 AR 拮抗剂抑制
3、3D肿瘤球体模型揭示AR调节治疗的差异效应
与传统的二维贴壁培养相比,3D 肿瘤球体模型被认为能更准确地反映肿瘤的复杂性。为了更好地模拟肿瘤微环境,作者使用超低成本贴壁板,从亲本 C4-2 或 LNCaP 细胞生成 LNCaP 和 C4-2 细胞的 3D 肿瘤球体模型。该模型提供了一个更符合生理相关性的系统,用于评估雄激素受体(AR)调节治疗(包括 SAL、ENZ 和 ODM)相对于 DMSO 对照的影响。
对球体体积的定量评估显示,用 SAL、ENZ 或 ODM 处理后,LNCaP 和 C4-2 球体的肿瘤大小均显著减小(图 4A,4B)。这一减少与之前在二维培养系统中的观察结果一致,强调了这些药物在破坏肿瘤生长方面的有效性。
对关键 AR 调节基因和 lncRNA 的分子分析显示出治疗特异性的表达模式。ADAMTS9-AS2,一种先前被认为参与 AR 信号调节的 lncRNA,在用 SAL、ENZ 和 ODM 处理后,在 LNCaP 和 C4-2 球体中的表达均显著增加(图 4A,4B)。这种模式支持其作为 AR 通路中潜在共调节因子的作用,并表明 AR 拮抗剂不会完全使 AR 信号失活。
相反,AR 靶基因,包括 TMPRSS2、KLK3 和 FKBP5 的表达,表现出对治疗类型的明显依赖性。这些 AR 靶基因在用 SAL 处理后上调,但在用 ENZ 和 ODM 处理的两种细胞系中均下调(分别见图 4A 和 4B)。这些差异反应突出了 SAL 作为 AR 激动剂与 ENZ/ODM 作为 AR 拮抗剂的对比效应。
有趣的是,lncRNA PART1 在用 SAL、ENZ 或 ODM 处理的 LNCaP 和 C4-2 球体中均显示出持续下调(图4A,4B)。这一发现与其作为 AR 信号共抑制因子的作用一致,特别是在 AR 通路调节的条件下。
这些结果共同突出了 3D 球体模型捕捉肿瘤特异性基因表达对 AR 靶向治疗反应复杂性的能力。它们还进一步证明了 AR 信号与 lncRNA 调节之间的相互作用,强调了它们在前列腺癌治疗中的相关性。
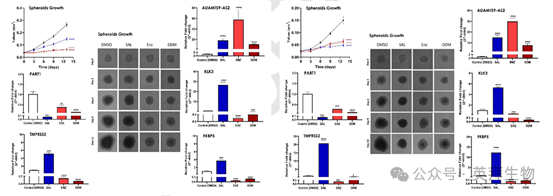
图 4A 和图 4B. SAL、ENZ 和 ODM 处理抑制 C4-2 和 LNCaP 细胞中的球体生长,并调节肿瘤球体中 ADAMTS9-AS2 和 PART1 的表达
4、C4-2细胞中AR对PART1和ADAMTS9-AS2基因座的差异招募
染色质免疫沉淀测序(ChIP-seq)分析表明,在 C4-2 细胞中,双氢睾酮(DHT)可使 AR 招募到 PART1(内含子)基因座(图 S4A)。此外,ChIP-seq 结果显示,在经 DHT 处理的 C4-2 细胞中,AR 选择性地招募到 ADAMTS9-AS2(内含子和外显子)基因座。因此,数据表明 PART1 是一种新型的 AR 靶基因,然而,它是一种非经典的 AR 靶基因,其表达被 SAL 处理抑制。
5、体外分析揭示SAL诱导ADAMTS9-AS2和PART1表达的显著变化
对前列腺切除术后未固定的原生患者前列腺癌标本进行 SAL 处理后的表达变化分析。作者检测到用 SAL 处理后,ADAMTS9-AS2和 PART1 的体外表达水平存在显著差异(图 5),证实了 SAL 对 LNCaP 和 C4-2 细胞中这些 lncRNA 的调节作用。
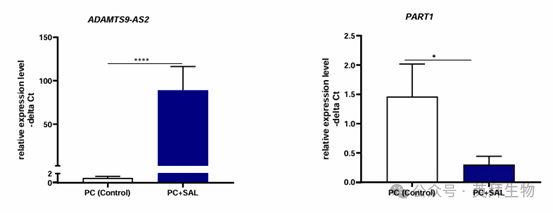
图 5. 在原生人类前列腺癌组织中,SAL 处理可控制ADAMTS9-AS2 和 PART1 lncRNA的表达
与 SAL 诱导 ADAMTS - AS2 表达类似,在经 SAL 处理的前列腺癌组织中,ADAMTS9-AS2 的体外表达显著升高(表达比值(95% 置信区间) = 54.2(16.9 - 175),P < 0.0001)。与 ADAMTS - AS2 相反,正如细胞系数据所预期的那样,经 SAL 处理的前列腺癌组织中 PART1 的体外表达降低(表达比值(95% 置信区间) = 0.12(0.02 - 0.77),P = 0.03)。此外,还检测到 PSA 水平与 BMI 之间存在显著相关性(相关系数 = 0.726,P = 0.027),PART1 表达与年龄之间存在显著相关性(相关系数 = -0.644,P = 0.049),以及 PART1 和 ADAMTS9-AS2 的表达水平之间存在显著相关性(相关系数 = 0.8,P = 0.10)。
6、lncRNA ADAMTS9-AS2是AR的共调节因子
获得的数据提出了一个假设,即 lncRNA ADAMTS9-AS2 与 AR 之间可能存在相互作用,影响AR 的转录活性。为此,通过小干扰RNA(siRNA)分别敲低 lncRNA ADAMTS9-AS2 和 PART1,并分析编码诊断标志物前列腺特异性抗原(PSA)的 KLK3、TMPRSS2 和 FKBP5(已知的直接 AR 靶基因)的表达。在两种细胞系中均验证了敲低效果(图 6a,6b),表明敲低有效。如预期的那样,AR 靶基因(FKBP5(b)、TMPRSS2(c)和 KLK3(d))被 SAL 上调。有趣的是,ADAMTS9-AS2的敲低降低了 SAL 诱导的 AR 靶基因 FKB5、TMPRSS 和 KLK3 的表达。这一观察结果表明,lncRNA ADAMTS9-AS2 可能作为 AR 的共激活因子发挥作用。因此,数据表明 lncRNA ADAMTS9-AS2 是 AR 信号通路的一部分。
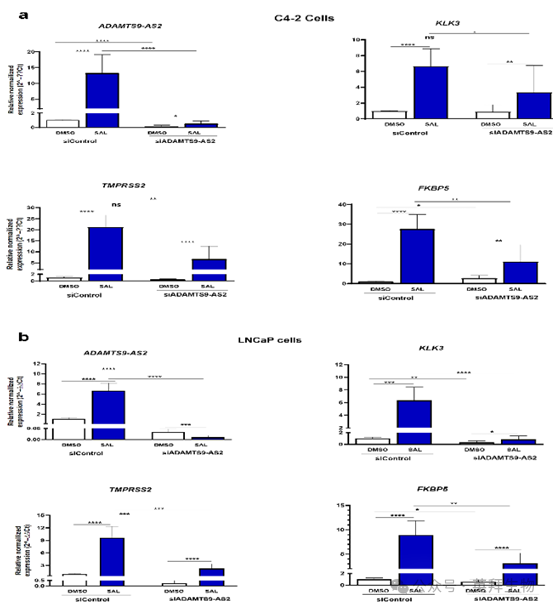
图 6a 和图 6b. ADAMTS9-AS2 的敲低降低了 AR 靶基因的表达
7、PART1 lncRNA敲低作为共抑制因子引发AR靶基因的过表达
同样,作者在 C4-2 和 LNCaP 细胞系中对 PART1 进行了敲低。与 ADAMTS-AS2 的敲低情况相反,长链非编码 RNA PART1 的表达受 SAL 抑制。敲低长链非编码 RNA PART1 会导致 AR 靶基因表达增强(图 7),这表明 PART1 是一种共抑制因子。与该观察结果一致的是,在患者前列腺腺癌数据集(GEPIA)中,PART1 的表达与 KLK3和 FKBP5呈显著正相关,与 TMPRSS2 的相关性较弱但也显著。因此,这些数据表明,长链非编码 RNA PART1可能作为一种共调节因子参与调节 AR12。
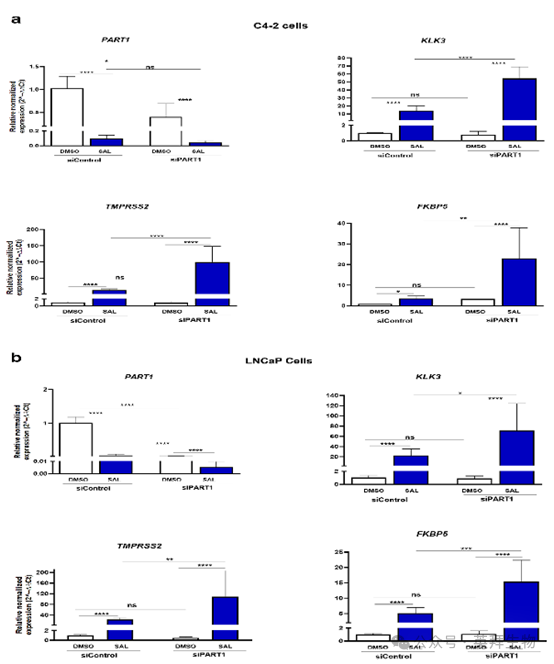
图 7a 和图 7b. PART1 的敲低
8、C4-2细胞中PART1和ADAMTS9-AS2的相互调节:对AR 信号传导和细胞衰老的影响
在通过 siRNA 介导敲低后,对 C4-2 细胞中 PART1 和 ADAMTS9-AS2 的表达分析揭示了一种动态调节的相互作用,这种相互作用影响着 AR 信号传导(图 8)。在没有 SAL 的情况下,siADAMTS9-AS2 敲低会导致 PART1 表达增加,PART1 作为共抑制因子抑制 AR 靶基因。相反,siPART1 敲低会增强 ADAMTS9-AS2 的表达,促进 AR 激活。这表明这两种长链非编码 RNA 之间存在相互作用。在 SAL 处理后,ADAMTS9-AS2 水平上升而 PART1 水平下降,进一步证实了一种相互的交互作用,即一种长链非编码 RNA 的增加对应着另一种的抑制。这种相互作用表明 AR 被 “超级激活”,从而驱动细胞衰老。
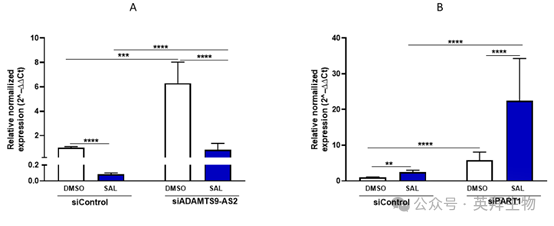
图 8 C4-2 细胞中经 siRNA 介导敲低和 SAL 处理后 PART1 和 ADAMTS9-AS2 长链非编码 RNA 的差异表达
9、RIP 测定突出了不同的富集模式:PART1被AR抗体显著富集,而ADAMTS9-AS2则没有
作者的结果揭示了在 LNCaP 细胞中使用 AR 抗体进行免疫沉淀时,沉淀物中存在独特的富集模式。值得注意的是,PART1 转录本在使用抗 AR 抗体免疫沉淀的物质中丰度大幅增加,这表明它与 AR 存在相互作用。与之形成鲜明对比的是,ADAMTS9-AS2 没有显示出显著的富集,这表明它与 AR 缺乏直接相互作用(图 9)。
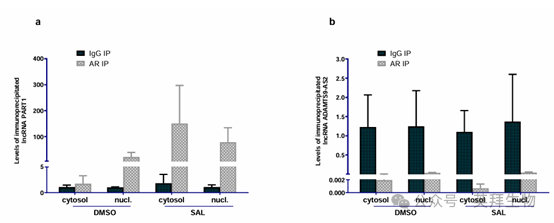
图 9. AR 与长链非编码 RNA PART1 结合。使用抗 AR 抗体通过 RIP 分析检测 RNA 结合蛋白(RBP)AR 与 PART1 和 ADAMTS9-AS2 长链非编码 RNA 的关联
10、siRNA介导的ADAMTS9-AS2敲低降低了SAL诱导细胞衰老的能力
用 SAL 处理前列腺癌细胞会诱导细胞衰老,这在体外处理的患者前列腺癌标本、前列腺癌 3D 肿瘤球体以及小鼠异种移植物中均有体现。由于长链非编码 RNA 调节 AR 信号传导,作者假设它们调节 SAL 介导的细胞衰老诱导过程。通过 siRNA 介导的敲低实验,作者的结果显示,靶向降低 ADAMTS9-AS2 的表达会显著减弱 SAL 在两种细胞系中诱导细胞衰老的能力(图 10)。这表明,被 SAL 上调的 ADAMTS9AS2 促进了 SAL 介导的细胞衰老。在研究 siRNA 介导的 PART1 敲低的影响时,作者观察到两种细胞系对 SAL 诱导的细胞衰老水平的细胞反应均有所降低。具体而言,在 C4-2 和 LNCaP 细胞中,降低 PART1 的表达会显著降低 SAL 诱导细胞衰老的能力。这种增强的敏感性意味着 PART1 和 ADAMTS9-AS2 在雄激素敏感和去势抵抗的前列腺癌细胞系中,在 AR 介导的衰老过程中均发挥着调节作用。
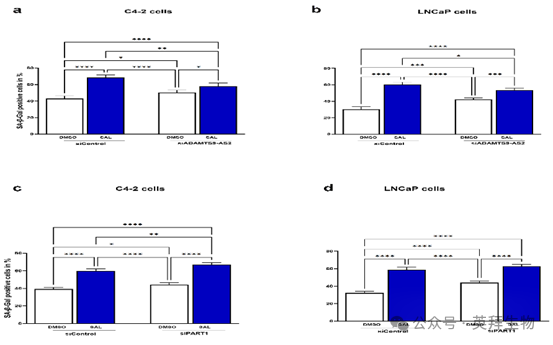
图 10. 敲低长链非编码 RNA ADAMTS9-AS2 会降低,而敲低 PART1 会增强 SAL 介导的细胞衰老
11、ADAMTS9-AS2和PART1敲低对C4-2 和LNCaP细胞中p15INK4B mRNA和蛋白水平的不同影响
在 C4-2 和 LNCaP 细胞中,分析了编码 p15INK4B 蛋白的 CDKN2B mRNA 水平,实验分为敲低 ADAMTS9-AS2 和未敲低的情况。结果表明,SAL 上调的 p15INK4B 的 mRNA 和蛋白水平均有所降低(图 11A - C),并且敲低 ADAMTS - AS2 会下调两种细胞系中 CDKN2B mRNA 和 p15INK4B 蛋白的水平(图 11C)。这一发现表明 ADAMTS9-AS2 在衰老诱导因子 p15INK4B 的表达中发挥调节作用。作者的数据也与衰老相关 β- 半乳糖苷酶(SA-β-gal)检测结果一致,该检测也显示敲低 ADAMTS9-AS2 后细胞衰老减少。因此,作者的观察结果进一步证实,敲低ADAMTS9-AS2 会降低 SAL 诱导衰老的能力,并表明 ADAMTS9-AS2 在一定程度上介导了 SAL 诱导的细胞衰老。
与 si - ADAMTS9-AS2 的情况相反,在 LNCaP 和 C4-2 前列腺癌细胞系中敲低PART1,会显著诱导p15INK4B 的 mRNA 和蛋白水平升高。这一观察结果强调了 PART1 在调节 p15INK4B 表达中的作用,表明 PART1 通常可能作为 p15INK4B 表达的抑制因子。作者还分析了另一种细胞周期抑制剂 p21,结果表明,与 ADAMTS9-AS2 不同,敲低长链非编码 RNA PART1 会增强 p21 蛋白水平,这表明 p21 也是 PART1 的作用靶点。有趣的是,在存在 SAL 的情况下,由于敲低两种长链非编码 RNA 会导致细胞衰老减少,p21 水平也会降低(图 11C)。
这些发现表明,SAL 介导的 PART1 下调是前列腺癌细胞中 SAL 介导的细胞衰老诱导过程的一部分。敲低 PART1 后 p15NK4B 的上调也表明了 PART1 促进前列腺癌进展,甚至可能增强其侵袭性的潜在机制。与该观察结果一致,来自患者前列腺癌样本的数据集显示 PART1 与 CDKN2B 之间存在显著的负相关。值得注意的是,在两种细胞系中,敲低 PART1 或 ADAMTS9-AS2 都会增强 AR 的蛋白水平(图 11C),这表明这两种长链非编码 RNA 可能通过调节内源性 AR 蛋白水平来调节 AR 活性16。
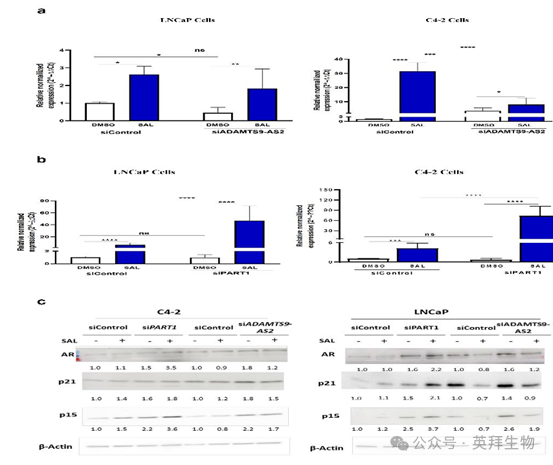
图 11. 细胞周期抑制剂和衰老标志物 p15 的水平受 SAL 以及敲低长链非编码 RNAs ADAMTS9-AS2 和 PART1 的影响
12、前列腺癌中ceRNA 网络的构建
在构建包含长链非编码 RNA(lncRNA)、微小 RNA(miRNA)和信使 RNA(mRNA)的竞争性内源性 RNA(ceRNA)网络时,研究人员聚焦于表达上调或下调的 miRNA。如果在 miRNA - mRNA 和 lncRNA - miRNA 相互作用对中,lncRNA 和 mRNA 与这些 miRNA 呈现相反的关联关系,那么它们就会被纳入研究范围。作者运用 miRcode来探究 lncRNA 和 miRNA 之间的联系。分析结果显示,在 9 种差异表达的 lncRNA 中,有 6 种可能靶向 55 种 miRNA 中的 17 种。作者使用 miRNAmeConverter 软件包(版本 1.30.0),将 miRNA 的名称转换为 miRbase v22 数据库中对应的名称。随后,利用 multimiR 软件包(版本 1.24.0)来识别 miRNA 的靶 mRNA。为了优化结果,作者剔除了那些不在差异表达 mRNA 范围内的 miRNA 靶向 mRNA。通过数据分析,作者确定了 13 种与前列腺癌相关的 miRNA,它们有可能靶向 266 种差异表达 mRNA 中的 107 种。此外,依据 ceRNA 网络理论,作者去除了 lncRNA 与 miRNA 之间、miRNA 与 mRNA 之间存在相反表达模式的相互作用。借助 Cytoscape 3.9 软件,作者构建了一个涵盖 3 种 lncRNA、13 种 miRNA 和 107 种 mRNA 的综合 ceRNA 网络。该网络包含 123 个节点和 124 条边。
作者使用 cytoHubba插件,基于最大团中心性、度中心性、介数中心性、径向中心性和离心率中心性,确定了 15 个枢纽基因。进一步分析这些枢纽基因的交集后,作者发现了一组包含 ADAMTS9 - AS2、hsa - miR - 10b - 5p、hsa - miR - 148a - 3p、hsa - miR - 183 - 5p、hsa - miR - 18b - 5p、hsa - miR - 199b - 5p、hsa - miR - 200a - 3p、hsa - miR - 203a - 3p、hsa - miR - 26b - 5p、hsa - miR - 301a - 3p、hsa - miR - 454 - 3p、hsa - miR - 7 - 5p 和 hsa - miR - 98 - 5p 的基因。作者还根据 ceRNA 网络中的 mRNA 构建了蛋白质 - 蛋白质相互作用(PPI)网络,并对该网络应用中心性分析。最后,通过共享这些中心性枢纽基因之间的相互作用,作者得到了 CAV1 和 FLNA 这两个枢纽 mRNA(图 11)。这些枢纽基因在网络中起着关键作用,对于理解它们在研究体系中的重要性至关重要。
对前列腺腺癌数据集的分析表明,CAV1 和 FLNA 的高表达水平与患者更高的生存概率呈正相关。这一发现凸显了这两个枢纽基因在前列腺癌病理生理学中的潜在重要性。该相关性与作者的生物信息学分析结果高度吻合,后者将 CAV1 和 FLNA 确定为调控网络的关键组成部分。这些结果表明,这些基因的上调可能通过调节参与肿瘤进展、细胞结构或转移行为的关键分子通路,为患者带来生存优势。不过,还需要进一步的实验验证,来阐明 CAV1 和 FLNA 的确切机制作用,并探索它们作为前列腺癌管理中的预后生物标志物或治疗靶点的潜力。
对前列腺腺癌患者中 CAV1和 FLNA 表达进行的 Kaplan - Meier 生存分析显示,它们与患者的预后存在显著关联。高 CAV1 表达(n = 334)与较低表达(n = 146)相比,患者的生存概率更高,这表明它有可能成为一种良好的预后标志物。同样,FLNA 的高表达与更好的总体生存率相关,这表明它在前列腺癌进展过程中具有保护作用。这些发现突出了 CAV1 和 FLNA 在前列腺腺癌预后方面的重要性。
13、ceRNA网络的关键轴
为了选择关键轴,作者通过枢纽基因之间的交集来获取基因。具体来说,在 ceRNA 网络中,作者利用枢纽基因之间的共享相互作用来选择 lncRNA 和 miRNA;对于 mRNA 的选择,作者则利用 ceRNA 网络中 mRNA 的 PPI 网络里基因枢纽之间的共享相互作用。包括 CAV1 和 FLNA 在内的 mRNA 位于主要 PPI 网络的最终基因枢纽中。这些关键轴包括 ADAMTS9 - AS2/hsa - miR - 7 - 5p/CAV1 和 ADAMTS9 - AS2/hsa - miR - 98 - 5p/FLNA(图 12)。
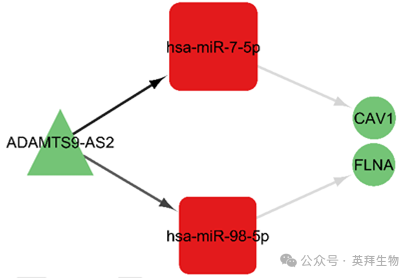
图 12. 基于枢纽基因的 ceRNA 网络关键轴
14、lncRNA - miRNA相互作用
此外,作者利用LncBase 数据库,探究了 miRNA 与 ADAMTS9 - AS2 和 PART1 之间的相互作用。作者实施了多种筛选条件来优化分析,例如通过组织筛选进行聚焦,将 miRNA 置信度筛选设置为 “高” 以确保 miRNA 相互作用的可靠性,并使用 “已验证” 筛选条件,仅纳入经过实验验证的相互作用。分析结果显示,PART1 与两种 miRNA,即 hsa - miR - 99a - 5p 和 hsa - miR - 150 - 5p 之间存在显著相互作用。此外,作者还发现 ADAMTS9 - AS2 与两种 miRNA,hsa - miR - 7 - 5p 和 hsa - miR - 98 - 5p 之间存在值得关注的相互作用。
15、lncRNA相互作用
通过 RNAinter 数据库,作者识别出 ADAMTS9 - AS2 和 PART1 与组蛋白修饰、转录因子(TFs)、蛋白质和 miRNA 的多种相互作用,这表明它们在染色质动态变化和转录调控中发挥着调节作用。ADAMTS9 - AS2 与活跃染色质标记,如H3K4me3(得分:0.8377)和 H3K27ac(0.7737),以及抑制性标记 H3K27me3(0.7579)都有很强的关联,这表明它在激活和抑制基因表达方面具有双重作用。它还与关键转录因子,包括 CTCF(0.6255)和 AR(0.4822),以及蛋白质 POLR2A(0.528)相互作用,这凸显了它作为染色质和转录调节因子的潜力。同样,PART1 与活跃组蛋白标记H3K4me3(0.6952)相关,并与转录因子,如 FOXA1(0.5984)和 AR(0.5479)相互作用,这表明它参与了转录调控,尤其是在激素驱动的通路中。这些发现突出了 lncRNA 在调节前列腺癌基因表达和细胞过程中的关键作用。
16、miR - 98 - 5p的表达上调,与ADAMTS9 - AS2存在显著相关性
为了验证生物信息学分析的结果,作者检测了临床样本中 miR - 98 - 5p 和 miR - 7 - 5p 的表达水平(图 13A)。与 NTAT 样本相比,这两种 miRNA 在 TT 样本中的表达均下调(表达比值(95% 置信区间)分别为0.033±0.76(0.011 - 0.097)和 29.65±0.6(12.85 - 68.4);miR - 98 - 5p 和 miR - 7 - 5p 的 P 值分别 < 0.0001 和 0.001)。另一方面,与 NTAT 样本相比,miR - 99a - 3p 和 miR - 150 - 5p 在 TT 样本中的表达也下调(表达比值(95% 置信区间)分别为19.22±0.46(10.1 - 36.8)和 0.32±0.56(0.15 - 0.71);miR - 99a - 3p 和 miR - 150 - 5p 的 P 值分别 < 0.0001 和 0.006)。生物信息学分析与患者数据集验证的结合,揭示了 ADAMTS9AS2 的一条新通路和一个新的 miRNA 网络。
为了进一步探究潜在的相互作用,作者进行了 Spearman 相关性分析。在前列腺癌患者的肿瘤组织样本中,miR - 99a - 3p 和 miR - 150 - 5p 的表达水平与 PART1 的相关性较弱(图 13B)。同样,miR - 7 - 5p 和 miR - 99 - 5p 与 ADAMTS9 - AS2 的相关性也较弱(图 13D)。尽管这些相关性未达到统计学显著性,但观察到的趋势,结合生物信息学预测和表达验证,表明存在一个涉及 ADAMTS9 - AS2 的潜在 miRNA 网络,值得进一步研究。
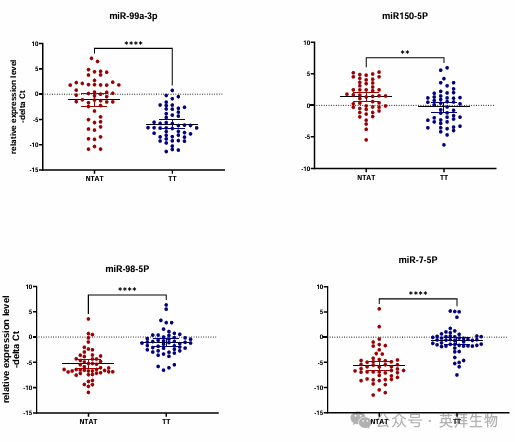 图 13A. 使用患者样本验证生物信息学预测结果
图 13A. 使用患者样本验证生物信息学预测结果
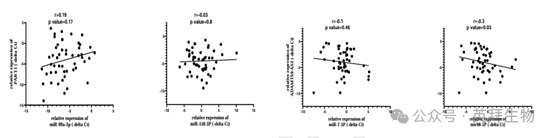
图 13B. 前列腺癌样本中 miRNA 与 lncRNA 的相关性分析
结论:
综上所述,作者发现了新的 AR 信号通路,这些通路涉及在前列腺癌肿瘤发生过程中被 SAL 反向调节的长链非编码 RNA。在双极雄激素治疗中使用的SAL,可诱导长链非编码 RNA ADAMTS9 - AS2 的表达,同时抑制长链非编码 RNA PART1 的表达。这两种长链非编码 RNA 均在 AR 信号通路中发挥作用,调控 SAL 介导的细胞衰老诱导过程。尽管它们受 SAL 的调控作用相反,但都参与了AR 信号传导,这表明在前列腺癌的AR 信号通路中存在新的途径和网络。这些发现有助于理解BAT所涉及的分子机制,为前列腺癌的治疗提供了潜在的新靶点和理论依据。不过,本研究存在一定的局限性,未来研究应注重体内验证,并采用患者来源的异种移植或类器官模型,以便更深入地探究这些发现的临床相关性。
实验方法:
差异表达分析、GO分析、通路富集分析、构建竞争性内源RNA(ceRNA)网络、相关性分析、生存分析、ChIP-seq、q-PCR、Western blot、RIP、细胞培养,siRNA敲低细胞转染、3D肿瘤球体模型构建、细胞衰老诱导实验、小鼠异种移植物实验
参考文献:
Taheri M, Schindler K, Baniahmad A. The LncRNAs PART1 and ADAMTS9-AS2 act in an Antithetic Manner on AR Signaling and Induction of Cellular Senescence in Prostate Cancer Cells. Int J Surg. 2025 Mar 26. doi: 10.1097/JS9.0000000000002334. Epub ahead of print. PMID: 40143747.