






免疫细胞竟靠“吃乳酸”增强战斗力?《Nature》子刊揭示抗感染代谢关键
训练有素的免疫力是指先天性免疫细胞的长期记忆。然而,人们对环境营养供应如何影响训练有素的免疫力知之甚少。本研究发现,生理碳源(PCSs)会影响葡萄糖对三羧酸(TCA)循环的贡献,并促进训练有素的单核细胞产生细胞因子。本研究证明,训练有素的单核细胞优先使用乳酸而不是葡萄糖作为 TCA 循环底物,乳酸代谢是训练有素的免疫细胞对细菌和真菌感染做出反应所必需的。除了对 TCA 循环的贡献外,内源性乳酸或外源性乳酸还通过调节组蛋白乳酰化支持训练有素的免疫。进一步的转录组分析、ATAC-seq和CUT&Tag-seq表明,乳酸盐以依赖组蛋白乳化的方式提高染色质的可及性。通过沉默乳酸脱氢酶 A(LDHA)抑制依赖乳酸的新陈代谢,损害以乳酸为燃料的 TCA 循环和组蛋白乳化。这些发现表明,乳酸是训练有素的免疫代谢和表观遗传程序的枢纽。该研究于2025年4月发表在《Nature Communications》,IF:14.7。
技术路线:

主要研究结果:
1. PCSs 在诱导训练有素的免疫过程中影响单核细胞对葡萄糖的利用
利用 13C 代谢示踪剂,作者研究了 PCSs 在训练免疫中对葡萄糖代谢的作用(图 1a)。质谱分析(MS)显示,13C-葡萄糖广泛标记了β-葡聚糖处理单核细胞中糖酵解和 TCA 循环的中间产物(图 1b-1d)。相比之下,在经 β-葡聚糖处理的单核细胞中存在生理浓度的 PCSs 时,13C-葡萄糖对 TCA 循环中间产物的贡献明显受到抑制(图 1b-1d)。耐人寻味的是,生理浓度下 PCS 的存在会引导葡萄糖流向 PPP 和丝氨酸合成途径(SSP)(图 1e 和 1d)。由 TCA 催化产生的代谢物,如谷氨酸和天门冬氨酸也得到了类似的结果(图 1g)。结果显示,βOHB 和乳酸通过柠檬酸合成酶(M + 2 柠檬酸)以 2 碳进入的方式为 TCA 循环提供碳,并在β-葡聚糖处理的单核细胞中贯穿整个循环(图 1h)。值得注意的是,与葡萄糖浓度(5 毫摩尔)相比,乳酸盐浓度(1 毫摩尔)更低,训练有素的单核细胞更愿意使用 PCS 而不是葡萄糖进行 TCA 循环代谢(图 1h)。综上所述,这些研究结果表明,在训练有素的免疫过程中,PCS 降低了13C 葡萄糖对 TCA 循环的贡献,并促使 13C 葡萄糖从糖酵解转向 PPP 和 SSP。
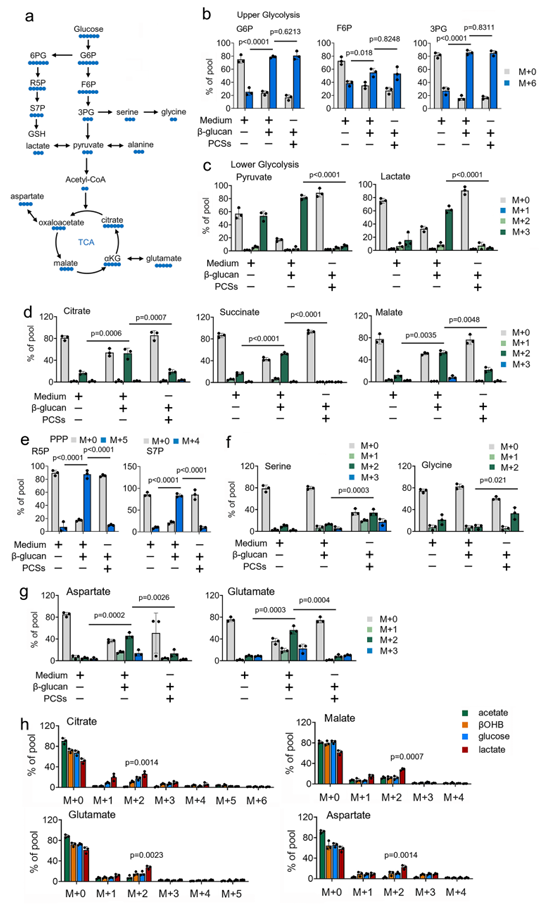
图1. PCS 影响葡萄糖在训练有素的免疫系统中的利用
2. 乳酸激活训练有素的免疫
由于在上述 PCSs 中,乳酸对 TCA 循环的调控作用最为显著(图 1h),作者接下来研究了乳酸在白僵菌感染时对训练有素的免疫系统的作用(图 2a)。由于乳酸脱氢酶(Ldh)通过使用 NAD+/NADH 作为辅助因子来相互转化乳酸和丙酮酸,作者提问LDHA 是否在训练有素的免疫中发挥作用。首先,采用他莫昔芬诱导的LDHA基因敲除小鼠(LDHA-/-)。为研究乳酸盐对训练单核细胞功能的作用,用β-葡聚糖训练小鼠,然后用白僵菌挑战小鼠(图 2a)。不出所料,训练过的β-葡聚糖 Wt 小鼠会诱导产生 IL-6 和 TNF-α,而 LDHA-/- 小鼠则不会(图 2b)。肺组织的组织学分析表明,在随后的感染过程中,事先进行β-葡聚糖处理可限制肺损伤(图 2c)。与接受过β-葡聚糖训练的Wt小鼠相比,LDHA-/-小鼠的肺损伤更为严重(图2c)。因此,LDHA-/-小鼠极易受到白僵菌诱导的致死影响,同时肺部和肾脏组织中的细菌负荷也较高(图 2d 和 2e)。
此外,作者还研究了乳酸盐在应对白僵菌感染时对训练有素的免疫力的作用。为排除乳酸盐通过改变酸度来影响训练有素的免疫力的可能性,在本手稿的所有体外实验中,培养基的 pH 值都被调整到相同水平。乳酸盐的存在增加白僵菌诱导的 IL-6 和 TNF-α水平,同时降低肾脏和肺组织中的真菌负担(图 2f-2h)。令人震惊的是,乳酸盐增加了训练有素的单核细胞对致命的全身性白僵菌感染的保护性反应(图 2i)。这些研究结果表明,乳酸对于训练有素的免疫系统对白茨球菌感染的保护性反应是不可或缺的。
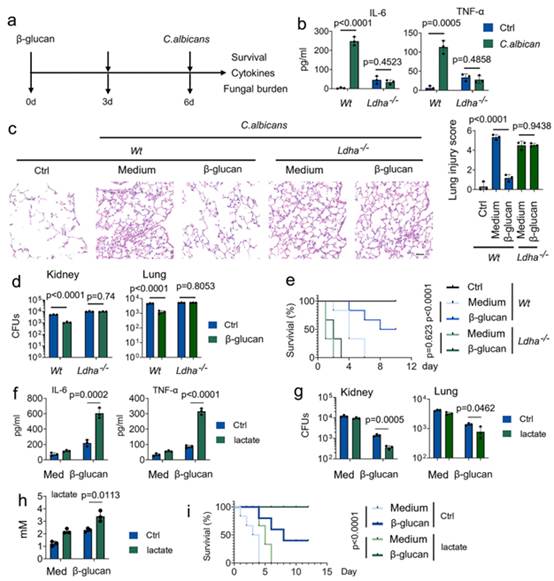
图2. 乳酸调控训练有素的免疫
3. 乳酸是诱导训练免疫过程中的一种生理燃料
为评估葡萄糖和乳酸盐在 TCA 循环代谢中的竞争情况,用 13C 葡萄糖和浓度逐渐增加的未标记乳酸盐培养处理过的单核细胞。即使在生理葡萄糖浓度(5 mM)下,β-葡聚糖处理过的单核细胞也会使用低浓度乳酸(1-2 mM)作为 TCA 循环中间产物的碳源(图 3a-3d)。相比之下,低浓度乳酸盐(1 mM)引导葡萄糖(5 mM)从 OxPhos 流向 SSP 和 PPP(图 3a-3f)。在乳酸盐处理的单核细胞中,葡萄糖衍生的大量标记 SAM(包括 m + 5(通过 PPP)和 m + 6-9(通过 PPP 和 SSP))显著增加(图 3g 和 3h),表明葡萄糖在乳酸盐存在的情况下被用于生物合成。由于乳酸有助于 TCA 循环代谢,接下来研究了乳酸对训练有素的单核细胞生物能的作用。通过使用以线粒体(Mito,mtAT1.03)为靶点的基于福斯特共振能量转移(FRET)的 ATP 传感器,发现乳酸盐的存在或β-葡聚糖的处理增强 ATP 的产生(图 3i)。同时处理β-葡聚糖和乳酸盐可显著协同诱导 ATP 的产生(图 3i)。与 13C 代谢物数据一致,乳酸增加了β-葡聚糖诱导的乙酰-CoA、柠檬酸盐和苹果酸盐的水平(图 3j)。最后研究了葡萄糖在β-葡聚糖处理过程中对乳酸促成的 TCA 循环代谢的作用。如图 3k 所示,在含有葡萄糖(1-10 mM)的培养基中,低乳酸浓度(1 mM)仍然有助于β-葡聚糖处理过程中的 TCA 循环中间产物。当葡萄糖浓度比乳酸浓度高 25 倍时,13C-乳酸不能为 TCA 循环提供碳(图 3k)。这些发现表明,在生理条件下,即使有葡萄糖,乳酸也是训练有素的单核细胞的 TCA 循环底物。
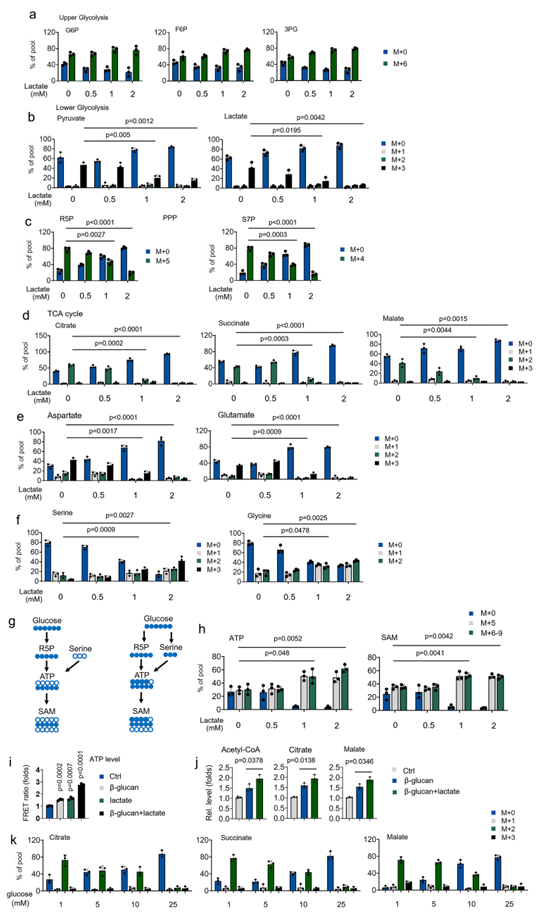
图3. 乳酸是诱导训练免疫过程中的一种生理燃料
4. 乳酸通过 LDHA 增强训练有素的免疫力
接下来研究 LDHA 在训练免疫中对乳酸生物能功能的作用。如图 4a 和 4b 所示,在 LDHA-/- 单核细胞中观察到 13C 乳酸衍生的柠檬酸盐、琥珀酸盐、苹果酸盐、谷氨酸盐和天门冬氨酸盐的丰度在β-葡聚糖处理后有所下降。13C 乳酸也有助于乙酰-CoA 相关代谢物的产生,包括 Ac-肉碱和 Ac-丙氨酸,这些代谢物存在于经 β-葡聚糖处理的 Wt 单核细胞中,但不存在于经 β-葡聚糖处理的 LDHA-/- 单核细胞中(图 4c)。通过使用线粒体 ATP 探针 mtAT,β-葡聚糖处理可提高 Wt 单核细胞的 ATP 产量,但 LDHA-/- 单核细胞经β-葡聚糖处理后的 ATP 产量则不会提高(图 4d)。同样,LDHA 基因敲除抑制β-葡聚糖诱导的乙酰-CoA、柠檬酸盐和苹果酸盐的水平(图 4e)。接下来,通过注入 13C 代谢物来探索宿主是否将乳酸作为体内训练免疫的燃料。在体内系统中,完全标记的(M + 3)13C-乳酸盐在血清和肺中富集了约 80%(图 4f 和 4g)。与体内外数据一致的是,输注的 13C 乳酸在体内很容易标记出 TCA 循环衍生的代谢物,这与β-葡聚糖处理 Wt 小鼠的结果一致,但在 LDHA-/- 小鼠体内则没有(图 4h 和 4i)。这些研究结果表明,通过 LDHA 处理乳酸是训练免疫的一个关键代谢节点。
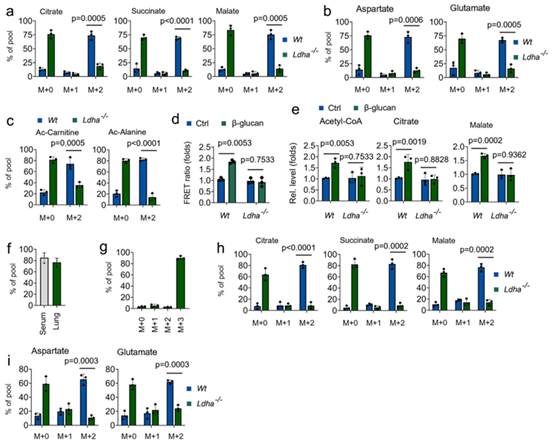
图4. Ldha 可影响新陈代谢和训练有素的免疫功能
5. 在诱导训练免疫过程中,乳酸盐通过 LDHA 调节单核细胞中组蛋白的乳化作用
先前的研究表明,赖氨酸乳酰化(Kla)是一种新型组蛋白标记,可被乳酸盐刺激;作者推测β-葡聚糖调节的高水平乳酸盐对训练免疫中组蛋白乳酰化的影响。免疫印迹分析表明,组蛋白 Kla 水平升高,但组蛋白乙酰化(Kac)水平轻度升高,且在β-葡聚糖处理的单核细胞中呈时间依赖性(图 5a)。组蛋白 Kla 水平的升高早在β-葡聚糖处理后 6 小时就被检测到(图 5a)。耐人寻味的是,高组蛋白 Kla 水平从第 1 天一直维持到第 9 天(图 5b)。不出所料,β-葡聚糖挑战以剂量依赖性方式诱导组蛋白 Kla 水平,并轻度诱导 Kac 水平(图 5c)。因此,使用 13C 葡萄糖或 13C 乳酸盐进行代谢标记实验,然后进行 MS/MS 分析,结果表明,β-葡聚糖可直接促进葡萄糖或外源乳酸盐的组蛋白乳化作用(图 5d-5g)。此外,β-葡聚糖处理增加 Wt 单核细胞中组蛋白 Kla 的水平,而 LDHA-/- 单核细胞中的组蛋白 Kla 水平却没有增加(图 5h)。这些研究结果表明,细胞内乳酸水平过高会导致免疫调节组蛋白乳酸化。
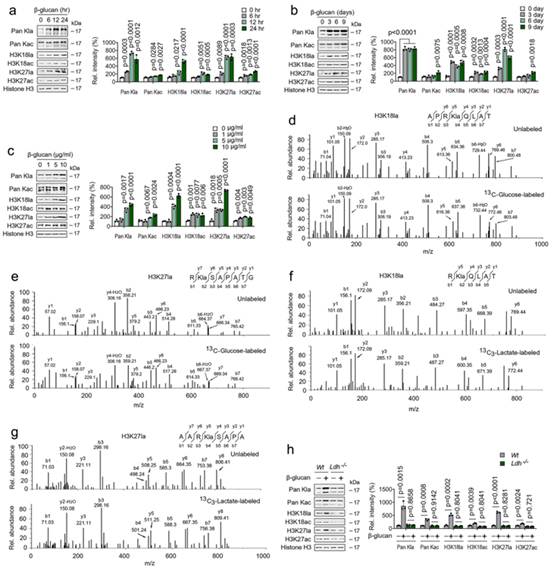
图5. β-葡聚糖通过 Ldha 调节组蛋白乳酸化
6. 在训练有素的免疫诱导过程中,乳酸通过组蛋白乳化作用维持单核细胞中染色质的可及性
为研究乳酸盐的产生对训练有素的免疫力的机械影响,利用高通量测序(ATAC-seq)分析法(Assay for Transposase-Accessible Chromatin with high throughput sequencing)比较了Wt单核细胞、Wt单核细胞和LDHA-/-训练有素的单核细胞之间染色质区域的可及性变化。确定了 2,848 个染色质区域出现可及性变化(图 6a)。为评估乳酸对转录的功能性影响,对 Wt 单核细胞、Wt 单核细胞和 LDHA-/- 训练有素的单核细胞进行生物重复的 RNA 序列分析(图 6a)。首先探讨染色质可及性变化与基因表达之间的关系。将每个差异可及区域(DARs)分配到与其最近的转录起始位点,并检测基因表达差异。观察到染色质可及性的变化与邻近基因的表达之间存在明显的相关性(图 6b)。接下来确定了在 Wt 单核细胞、Wt 和 LDHA-/- 训练单核细胞中上调的 836 个基因,这些基因也标记有染色质可及性的改变(图 6c)。对这些基因进行的基因本体(GO)生物过程分析显示了生物通路的显著富集,包括免疫反应、乳酸代谢和 HIF 信号转导(图 6d)。选择H3赖氨酸27(H3K27la)(该赖氨酸残基也有乙酰化(H3K27ac)),利用新型的全基因组免疫系留测定法CUT&Tag来阐明组蛋白乳酰化在训练有素的免疫中的潜在功能意义。全基因组分布分析表明,H3K27la 和 H3K27ac 组蛋白修饰主要位于启动子区域(图 6e)。正如预期的那样,在 Wt 训练的单核细胞中,H3K27la 峰在转录起始位点(TSSs)附近增加,而在 LDHA-/- 训练的单核细胞中则没有增加(图 6f)。比较H3K27la标记基因在Wt非训练单核细胞、Wt和LDHA-/-训练单核细胞中的表达。在三组共 7547 个共有峰中,发现有 248 个峰发生显著变化(图 6g)。GO通路分析显示,炎症反应、乳酸代谢和细胞粘附等生物过程与ATAC-seq数据一致(图6h)。为检验 H3K27la 和 H3K27ac 在训练有素的单核细胞功能中的作用,将 ATAC-seq 数据集和 CUT&Tag 数据集整合到整合基因组学浏览器(Integrative Genomics Viewer)中,以检查它们在基因上的结合情况。在 Wt 训练的单核细胞中,包括 IL-6、TNF-α、PSGL 和 ICAM-1 在内的炎症因子和细胞粘附分子的关键基因启动子上的 H3K27la 峰高度富集,而在 LDHA-/- 训练的单核细胞中则没有(图 6i、6j)。然后使用染色质免疫沉淀(ChIP)-qPCR 试验评估 H3K27la 和 H3K27ac 对稳定效应基因表达的影响。事实上,在Wt-训练的单核细胞中,IL-6、TNF-α、PSGL和ICAM-1启动子区域的H3K27la招募增加,但在LDHA-/-训练的单核细胞中却没有增加(图6k)。与 Wt 非训练型单核细胞相比,Wt 训练型单核细胞中 IL-6、TNF-α、PSGL 和 ICAM-1 启动子区域的 H3K27ac 募集仅存在微小差异(图 6l)。这些发现表明,内在乳酸盐的产生改变了训练有素的单核细胞中炎症因子和细胞粘附分子启动子上乙酰化依赖性表观遗传编程。
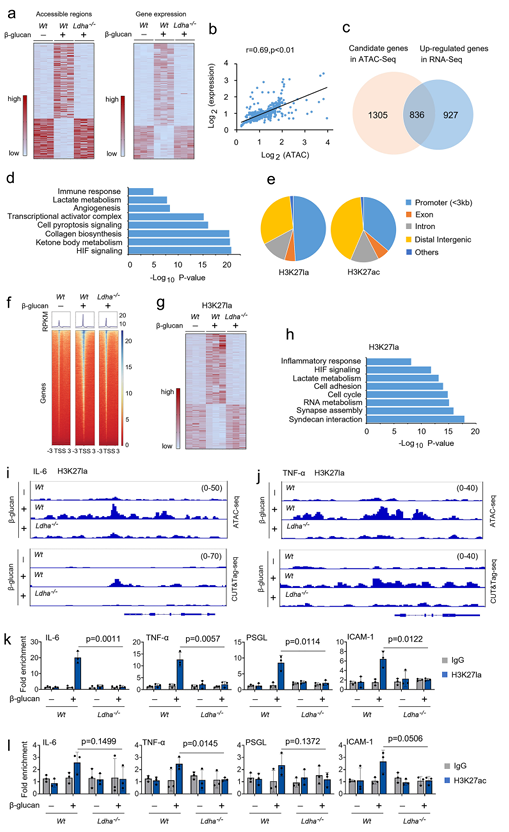
图6. 在诱导训练免疫的过程中,乳酸通过 Ldha 调节单核细胞中组蛋白的乳酸化
7. 外源性乳酸盐通过促进 TCA 循环和组蛋白乳化作用,支持训练有素的免疫力
作者的数据显示,β-葡聚糖能温和地诱导组蛋白乙酰化。因此推测,乳酸被吸收后很容易通过 LDHA 转化为丙酮酸,然后在丙酮酸脱氢酶(PDHA)和 ATP 柠檬酸裂解酶(ACLY)的两种酶催化反应中分别转化为细胞质乙酰-CoA33(图 7a)。为此,比较了葡萄糖和乳酸对乙酰-CoA 生成的作用。如图 7b 所示,高葡萄糖(HG)和低葡萄糖(LG)都不影响β-葡聚糖调节乙酰-CoA 的生成,但乳酸盐处理可诱导β-葡聚糖调节乙酰-CoA 的生成,表明β-葡聚糖通过乳酸盐促进乙酰-CoA 的生成。但葡萄糖却不能(图 7b)。为剖析乙酰-CoA生成所涉及的乳酸代谢的关键步骤,分别采用LDHA抑制剂(草氨酸钠,Oxa)、丙酮酸转运体抑制剂(UK5099)和ACLY抑制剂(BMS303141,BMS)等一系列针对LDHA、PDHA和ACLY的药理学方法(图7a)。有趣的是,Oxa、UK5099 和 BMS 可抑制β-葡聚糖诱导的乙酰-CoA 生成(图 7c)。以乙酰-CoA 的同位素为重点的 LC/MS 分析表明,乳酸碳标记大部分乙酰-CoA。m + 2 乙酰-CoA同位素显示了来自乳酸碳的最重要的标记模式(图 7d 和 7e)。为明确乳酸促进组蛋白乙酰化的途径,假设这涉及氧化代谢,尤其是 ACLY。令人吃惊的是,BMS能抵消β-葡聚糖介导的组蛋白乙酰化,但不能抵消组蛋白乳酸化(图 7f)。令人惊讶的是,ACLY抑制剂并不影响β-葡聚糖介导的IL-6和TNF-α的产生,也不影响体外和体内肺和肾组织中的细菌负荷(图7g-7i)。此外,BMS 处理抑制 H3K27ac 和 H3K18ac,但不能抑制 H3K27la 和 H3K18la 在 IL-6、TNF-α 和 PSGL 启动子区域的招募(图 7j-7m)。这些结果表明,乳酸是乙酰-CoA 生成的重要底物。与乳酸介导的组蛋白乙酰化相比,乳酸介导的组蛋白乳酸化在训练免疫中起着主导作用。
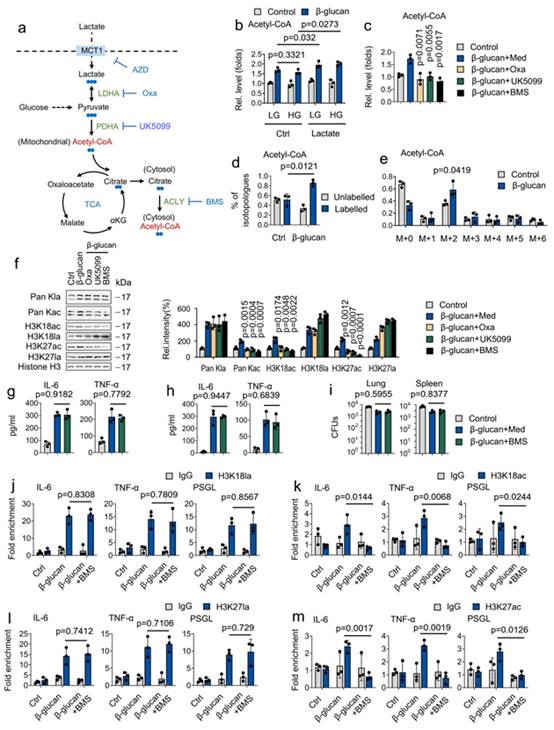
图7. 外源性乳酸盐通过促进 TCA 循环和组蛋白乳化作用,支持训练有素的免疫
结论
综上所述,本研究证明乳酸是训练有素的免疫细胞的生理燃料,比葡萄糖更受青睐,并能调节训练有素的免疫细胞的细胞因子产生。从机理上讲,一些固有的乳酸盐功能是乙酰-CoA 合成的重要底物,并为 TCA 循环提供燃料。另一种固有乳酸则进入细胞核,通过组蛋白乳酸化作用调节细胞因子的产生。这些数据确定了训练免疫中的主要代谢物(乳酸),有助于我们了解先天性免疫记忆,并开辟新的治疗途径。
实验方法:
单核细胞和外周血单核细胞分离,细胞培养,Western blot,细胞因子测量,ChIP-seq,CUT&Tag seq,ATAC seq,细菌培养,ChIP-qPCR,质谱,流式细胞量化线粒体 ATP 水平
参考文献
Cai H, Chen X, Liu Y, Chen Y, Zhong G, Chen X, Rong S, Zeng H, Zhang L, Li Z, Liao A, Zeng X, Xiong W, Guo C, Zhu Y, Deng KQ, Ren H, Yan H, Cai Z, Xu K, Zhou L, Lu Z, Wang F, Liu S. Lactate activates trained immunity by fueling the tricarboxylic acid cycle and regulating histone lactylation. Nat Commun. 2025 Apr 4;16(1):3230. doi: 10.1038/s41467-025-58563-2. PMID: 40185732; PMCID: PMC11971257.