






天然多酚激活并增强GPX4以减轻淀粉样蛋白β诱导的阿尔茨海默病铁死亡

阿尔茨海默病(AD)是一种常见的神经退行性疾病,也是最常见的痴呆形式,其特征是记忆力和认知功能的进行性下降。尽管研究取得了重大进展,但对其病因的了解仍然有限。铁死亡是一种铁调节的细胞死亡方式,近年来因其参与多种疾病的发生发展而在病因学研究和治疗方面受到关注。细胞内铁水平是铁死亡的关键因素。铁还作为特异性脂氧合酶的辅因子促进酶促脂质过氧化。然而,启动铁死亡的铁浓度阈值仍不明确。线粒体功能障碍、脂质过氧化以及抗氧化酶谷胱甘肽过氧化物酶4(GPX4)活性和水平的降低与铁死亡相关。虽然铁死亡正在被探索作为一种消灭肿瘤的治疗途径,但已发现它在多种神经系统疾病中发挥拮抗作用,包括AD。目前还需要开发同时靶向铁死亡和AD的多功能分子,并协同增强和激活包括GPX4在内的天然抗氧化机制,以减轻AD中铁死亡的复杂病理。该研究发表在《Chemical Science》,IF:9.825。
技术路线

图1 机制图

主要研究结果
1. 多酚(PPs)的铁结合作用
铁死亡是一种铁依赖的细胞死亡方式,铁螯合剂被认为是一种有前景的铁死亡抑制剂。评估了一组主要含有儿茶酚和没食子酸酯部分的PPs的铁螯合能力。筛选的PPs包括没食子酸(GA)、表没食子儿茶素没食子酸酯(EGCG)、鞣酸(TA)、鞣花酸(EA)、染料木黄酮(GEN)、丁香酸(SA)、没食子酸甲酯(MeG)、4-羟基黄酮(4-HF)和2-羟基黄酮(2-HF)(图1B)。在HEPES缓冲液中使用紫外-可见吸收光谱测定了这些PPs与Fe3+的相互作用强度。在加入FeCl3后,PPs的紫外-可见吸收光谱显示吸光度增加,其中TA增加最大(图2A)。将数据拟合为Benesi-Hildebrand方程,以确定结合常数(KB)。TA含有5个没食子酸,表现出最高的铁螯合能力,其KB值为6.64 ×105 M−1,而EGCG和GA的KB值分别为5.1×105 M−1和4.2×103 M−1(图2B)。研究者发现MeG,GEN,EA和2-HF的结合常数远低于TA。
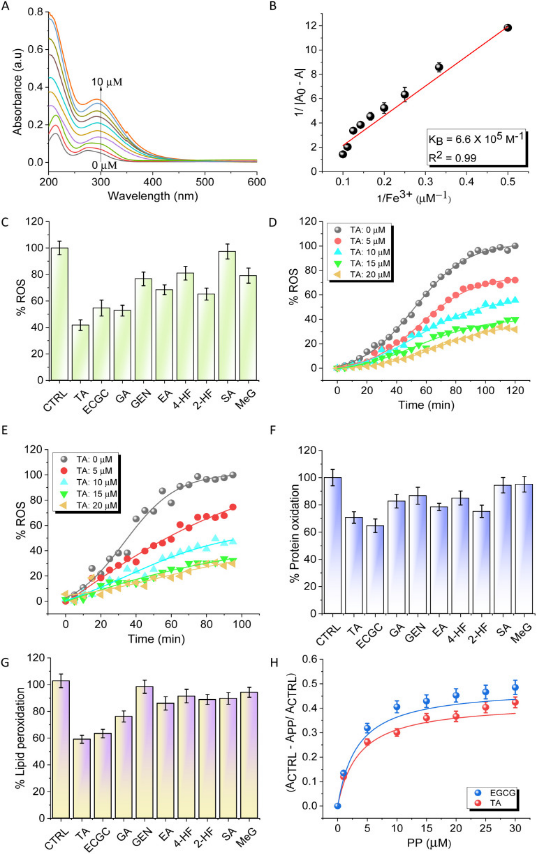
图2 PPs的铁螯合和抗氧化研究
2. 抗氧化作用和抑制脂质过氧化
采用α-二苯基-β-苦酰基肼(DPPH)、ABTS 和抗坏血酸铁实验评价强铁螯合剂TA和EGCG以及GA的抗氧化活性。通过在PBS缓冲液和FeCl3中使用铁抗坏血酸测定来评估PPs对氧化还原沉默Fe3+和猝灭过量ROS的能力(图2C)。TA在452nm处以浓度依赖性的方式降低了发射,表明它能捕获Fe3+并稳定氧化还原休眠状态,并清除自由基(图2D)。在最高浓度20μM时,TA对ROS的猝灭率为80%。有趣的是,在Aβ42存在的情况下进行的模拟AD条件的类似实验表明,TA可以有效结合并阻止Fe3+氧化还原过程,从而猝灭ROS的产生(图2E)。研究者使用Fe2++H2O2氧化还原体系来评估PPs对牛血清白蛋白氧化的保护作用,该体系在原位产生ROS,并模拟类似铁死亡的铁介导的氧化应激环境。TA和EGCG处理分别减少了30%和35%的蛋白质氧化(图2F)。
接下来,研究者使用α-磷脂酰胆碱作为模型脂质和Fe2++H2O2氧化还原体系进行了脂质过氧化测定,以测量铁死亡条件下的脂质过氧化程度。以Fe2++H2O2对α-磷脂酰胆碱的过氧化作用作为对照,发现TA和EGCG对脂质过氧化作用的抑制率分别为42%和40%(图2G)。在TA和EGCG存在的情况下,脂质过氧化的浓度依赖性图显示了它们出色的过氧化抑制活性,这可以归因于它们的协同铁螯合和自由基捕获机制(图2H)。
3. 铁非依赖性和依赖性Aβ42和tau蛋白聚集的调节
研究者评估了PPs调节Aβ42铁非依赖性和依赖性聚集倾向的能力聚集依赖性硫黄素T(ThT)荧光数据显示,在所有PPs中,EGCG和TA表现出优越的活性,在浓度为20μM时,聚集抑制率分别为44%和35%(图3A)。Aβ42的聚集滞后时间(Tlag)为9.2h,一级聚集速率常数(κ)为0.51 h−1。TA和EGCG浓度的增加增加了Tlag,降低了κ,表明Aβ42聚集显著延迟(图3B)。当Aβ42∶PP为1∶5时,EGCG的抑制率(95%)明显高于TA(82%)。由于铁稳态在AD病因学中的关键作用,研究者研究了铁对Aβ42聚集的影响。与原始Aβ42相比,Fe3+ 1:1的Aβ42使Tlag下降至8.72 h,而κ从0.51 h−1增加至0.72 h−1(图3C)。在1:5比例下,TA和EGCG的Tlag值分别为10.12 h和10.9 h,κ值分别为0.20 h−1和0.17 h−1(图3D)。
使用ThT荧光动力学监测PPs调节花生四烯酸诱导的tau聚集的能力(图3E)。EGCG和TA在浓度为20μM时,对细胞的聚集抑制率分别为53%和46%。TA和EGCG表现出浓度依赖性的活性,在1:5的比例下,抑制率最高,分别为70%和81%,在2.5小时之后,没有观察到ThT荧光进一步增加(图3F)。

图3 筛选PPs抑制Aβ的能力42并使用ThT测定监测tau聚集
4. GPX4的激活及其RSL3诱导的抑制的逆转
GPX4缺乏增加了细胞对铁死亡的易感性。GPX4有两个不同的位点:底物结合位点和变构位点。与抑制剂位点结合的小分子可作为抑制剂,而与激活位点结合预计可增强GPX4活性(图4A)。TA与GPX4的激活位点结合,而EGCG与GPX4的激活位点或抑制位点不一致。研究者对GPX4抑制剂RSL3和GPX4激活剂PKUMDL-LC-102进行了对接研究(图4A)。结果表明,RSL3与抑制位点结合,而PKUMDL-LC-102与激活位点结合,类似于TA。在变构激活位点,GPX4激活剂PKUMDL-LC-102被酸性残基D21和D23以及极性残基V98、F100和M102包围(图4A)。
使用GPX4抑制剂筛选试剂盒评估TA对GPX4活性的影响。在该实验中,研究者利用GPX4对NADPH的消耗来监测酶的活性。GPX4与RSL3孵育60分钟可抑制GPX4活性22%。然而,在20μM的浓度下,TA和EGCG能够分别逆转RSL3诱导的抑制作用12.3%和9.5%(图4B)。TA通过结合GPX4的激活位点来增加GPX4的活性。通过监测RSL3对GPX4活性的浓度依赖性抑制,研究者计算出了RSL3的半最大抑制浓度(IC50)值,结果为17.17±0.17μM(图4C,D)。RSL3的IC50值是在TA存在的情况下独立计算的。在反应混合物中加入20 μM的TA将RSL3的IC50值从17.17±0.17μM调节到29.98±1.09μM,表明TA可以降低RSL3诱导的GPX4抑制(图4E,F)。EGCG在RSL3诱导的GPX4抑制后显示出相对较低的恢复内在酶活性的能力,将IC50值改变为21.01±0.47μM(图5B)。对接结果表明TA具有与GPX4激活位点结合的能力。再加上TA可以有效逆转RSL3诱导的GPX4抑制,因此研究者研究TA是否可以直接作为GPX4激活剂发挥作用。研究者的数据表明,用TA处理GPX4导致酶活性的浓度依赖性增加。在100 μM TA浓度下,观察到GPX4活性增强了19%(图4G)。这些结果表明,TA不仅可以从RSL3诱导的抑制中恢复GPX4的活性,而且还可以增强其内在活性。所有体外实验的综合分析确定TA是最有希望的抗铁死亡候选药物。这主要是因为它能够激活GPX4并伴有多功能活性。相比之下,EGCG显示出更好的抗淀粉样蛋白和tau蛋白特性。
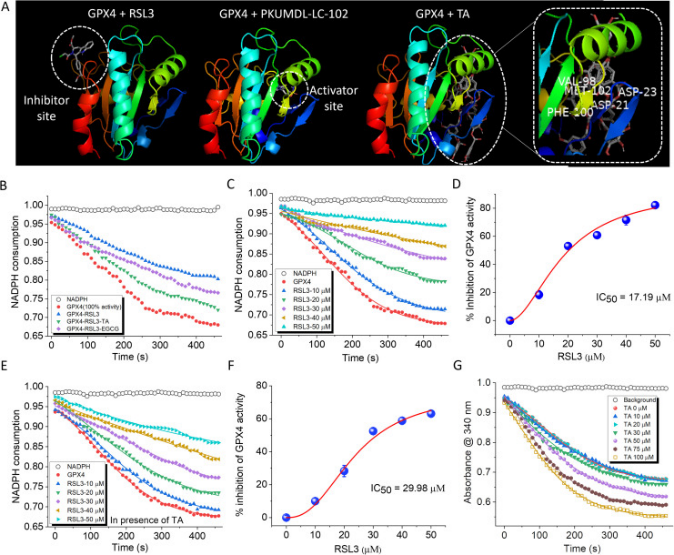
图4 GPX4在小分子与抑制剂和激活剂位点结合的情况下的酶活性
5. 挽救铁死亡诱导的细胞死亡
培养48h后,采用噻唑蓝(MTT)法检测铅PPs对SH-SY5Y神经元细胞活力的影响。为了确定TA和EGCG抑制铁死亡的能力,研究者将SH-SY5Y细胞分为RSL3单独处理组和TA和EGCG联合处理组。培养48h后,RSL3处理组细胞存活率为46%,而未处理组细胞存活率为100%。值得注意的是,TA和EGCG以浓度依赖性的方式拯救神经元细胞,在浓度为20μM时,细胞存活率分别达到84%和62%(图5A,B)。研究者进行了TA与Ferrostatin-1和DFO的比较研究,以研究RSL3诱导的铁死亡的拯救能力。Ferrostatin-1在10μM的低浓度下几乎能100%抑制铁死亡。TA挽救RSL3诱导的细胞死亡的效果与较低浓度的Ferrostatin-1相当。另一方面,尽管DFO具有比TA更强的铁螯合能力,但在一定浓度下,DFO并未表现出显著的拯救铁死亡损伤的作用。采用免疫荧光法检测RSL3单独处理以及TA和EGCG联合处理后细胞中GPX4的水平。与健康细胞相比,RSL3处理24 h可将GPX4水平降低至83%。值得注意的是,TA和EGCG处理使RSL3处理的细胞中的GPX4水平分别恢复至96%和91%,表明细胞从铁死亡中恢复(图5D)。此外,为了验证这些结果,研究者使用了蛋白质印迹分析来定量GPX4的水平。与对照细胞相比,RSL3处理后GPX4水平降低至73%(图5E、F)。值得注意的是,经TA治疗后GPX4水平完全恢复。
LIP升高是铁死亡的标志,可催化脂质过氧化并导致细胞死亡。用钙黄绿素-乙酰氧基甲酯染色检测RSL3单独处理、TA和EGCG联合处理的细胞LIP。钙黄绿素-乙酰氧基甲酯在细胞内被酯酶裂解,在细胞质中产生钙黄绿素荧光团,表现出高荧光。然而,当钙黄绿素与LIP螯合时,其荧光被猝灭,从而可以估算游离LIP水平。与健康细胞相比,RSL3处理的细胞显示细胞内Fe水平增加,相应的钙黄绿素荧光降低至67%(图5G-K)。值得注意的是,50μM TA和EGCG处理降低了LIP水平,导致钙黄绿素荧光分别增加约95%和100%,与对照细胞相似(图5L)。这种作用呈剂量依赖性,表明TA和EGCG能够隔离游离Fe并使其失活。铁死亡触发脂质过氧化,增加ROS生成,导致细胞内线粒体ROS水平升高和线粒体膜电位(MMP)下降,导致线粒体碎片化。研究者使用MitoSOX探针监测细胞中线粒体ROS水平。RSL3处理使线粒体ROS水平增加到148%,与健康细胞相比。相反,TA和EGCG分别将RSL3处理的细胞中的ROS水平降低至95%和91%,表明它们有能力预防铁死亡ROS介导的线粒体损伤(图5M)。通过探针检测MMP的变化,证实RSL3诱导的铁死亡和PP处理的逆转。RSL3处理细胞后,MMP下降至76%,与正常细胞比较差异有统计学意义。然而,TA和EGCG处理分别将MMP恢复至93%和95%,表明对功能性线粒体损伤有预防作用(图5N)。采用Mito-TG.45染色法检测TA和EGCG对铁死亡细胞线粒体结构损伤的保护作用采用活细胞荧光显微镜观察RSL3单独处理或TA和EGCG联合处理的细胞。值得注意的是,经TA和EGCG处理的细胞保持了健康的管状结构,支持这些PPs对铁死亡线粒体损伤的保护作用,ROS和MMP研究证明了这一点(图5R,S)。

图5 细胞研究表明TA和EGCG能够从铁死亡和Aβ中拯救神经元细胞42-诱导毒性
6. 从淀粉样蛋白诱导的铁死亡中拯救神经元细胞
TA和EGCG调节Aβ42聚集的有效性促使研究者研究它们拯救神经细胞免受Aβ42诱导的毒性的能力。分别用Aβ42、TA和EGCG处理SH-SY5Y细胞。Aβ42处理细胞30h后,细胞存活率为34.8%,与未处理的对照组相比,差异有统计学意义。然而,在PPs存在的情况下,研究者观察到Aβ42处理的神经元细胞以剂量依赖性的方式得到挽救,最高浓度的TA和EGCG分别使细胞存活率提高到63%和69%(图5T)。在AD的铁死亡背景下,铁加速毒性淀粉样蛋白(Aβ42-Fe)纤维的积累,产生活性氧,导致膜损伤和脂质过氧化。研究者使用MTT法检测TA和EGCG调节毒性Aβ42 + Fe聚集和拯救神经元细胞免于Aβ42 + Fe诱导的铁死亡的能力。Aβ42: Fe(1: 5)处理组细胞存活率为17%,对照组细胞存活率为100%。然而,用TA和EGCG处理后,Aβ42 + Fe处理的细胞存活率分别提高到74%和77%,呈剂量依赖性(图5U,V)。将细胞分为Aβ42 + Fe组和TA + EGCG组,DCFDA法检测细胞内ROS水平;Aβ42 + Fe(1:5)培养6h后,细胞内ROS由未处理组的31%增加至100%。然而,TA和EGCG处理后,Aβ42 + Fe处理细胞中的ROS水平分别降低至36%和38%,表明它们具有复合体和使Fe氧化还原失去活性、调节Aβ42 - Fe fenton型反应并最终阻止ROS生成的能力(图50)。这些细胞研究有力地支持TA和EGCG能够减轻RSL3诱导的铁死亡和Aβ42 - Fe相关毒性,使它们成为协同靶向铁死亡和AD的潜在候选药物。结合ThT研究,研究者评估了TA保护神经元细胞免受tau诱导的毒性的能力。在AA诱导的tau聚集物存在下,与健康细胞相比,细胞活力降低至47%(图5W)。TA可以剂量依赖性地增加细胞活力,将细胞活力提高到100%。
为了证实Aβ42与铁死亡之间的关系,研究者采用免疫荧光法检测Aβ42 + Fe单独孵育以及TA和EGCG共同孵育的SH-SY5Y神经元细胞株中GPX4的水平。与健康细胞相比,Aβ42 + Fe(1:5)处理细胞30小时显著降低GPX4水平至65%(图6)。据研究者所知,这是第一个在铁死亡背景下的神经细胞中Aβ42 + Fe和GPX4水平直接相关的报告,通过直接定量蛋白水平的免疫荧光测定确定。与未处理的对照细胞相比,TA显著增加了Aβ42 + Fe处理的细胞GPX4水平(147%),表明TA可以在AD病理条件下提高GPX4水平。
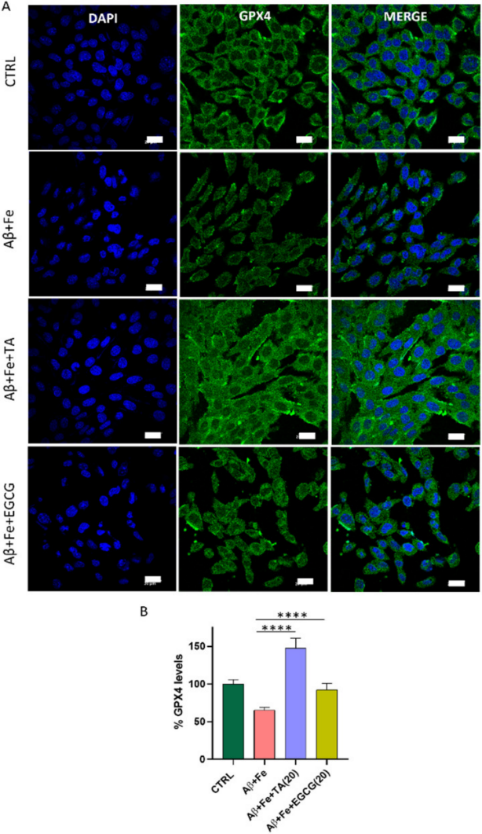
图6 通过免疫荧光(IF)测定进行蛋白质定量
这些发现促使研究者在神经元细胞中检测TA作为GPX4增强子的作用。研究者观察到,与未处理的细胞相比,TA处理的细胞内GPX4水平增加到192%(图7A,B)。TA不仅增强GPX4活性,而且增加其在细胞中的水平。蛋白质印迹检测GPX4蛋白表达水平。结果显示,与未处理的细胞相比,TA处理使GPX4水平增加到127%(图7C,D)。基于这些发现,研究者检测了抗氧化主要调节因子核因子e2相关因子2(Nrf2)的变化,已知Nrf2可直接和间接调节GPX4及其相关抗氧化蛋白46虽然有报道表明TA能够激活Nrf2通路,但研究者评估了在神经细胞中TA处理后Nrf2的激活。研究表明,TA处理导致SH-SY5Y细胞中Nrf2积累和核转位。与未处理的细胞相比,细胞内Nrf2水平增加到148%,并且使用TA处理后,来自细胞质的Nrf2蛋白发生了2倍的核易位(图7E-G)。因此,研究者推测TA可能通过Nrf2-GPX4轴提高GPX4水平。

图7 通过IF测定和蛋白质印迹进行蛋白质定量
结论
该研究发现天然的TA是一种高效的铁死亡抑制剂。它通过铁螯合、抑制脂质过氧化、拯救线粒体损伤和激活Nrf2-GPX4轴等铁死亡的所有主要途径发挥作用,从而提高它们的水平。同时,TA是一种有效的Aβ42和tau蛋白聚集的抑制剂,有效地调节剧毒铁诱导的Aβ42聚集,减少氧化应激,对抗AD患者的铁稳态失调。虽然GPX4蛋白的合成效率非常低且需要大量能量,但研究者证明了一种天然的多酚如何提供一种可行的解决方案,以协同调节铁死亡和AD的多方面毒性,而GPX4的激活是关键机制之一。这可能鼓励研究人员探索无毒的天然产物来解决问题,并强调GPX4在有效调节铁死亡以及阐明其在AD中的作用的重要性。
实验方法
ABTS抗氧化试验,铁抗坏血酸测定,硫黄素T(ThT)荧光检测,细胞培养和MTT法检测,ROS测定,神经细胞拯救实验,温滴定量热法(ITC)测量,蛋白质印迹,免疫荧光
参考文献
Baruah P, Moorthy H, Ramesh M, Padhi D, Govindaraju T. A natural polyphenol activates and enhances GPX4 to mitigate amyloid-β induced ferroptosis in Alzheimer's disease. Chem Sci. 2023 Aug 22;14(35):9427-9438. doi: 10.1039/d3sc02350h.