






高分肿瘤文章的剖析——计算模拟+单细胞转录组学+机制

胶质母细胞瘤在其早期阶段的进展仍然知之甚少。本研究中,作者在小鼠大脑中转移PDGFB和genetic barcodes,以启动胶质瘤发生,并能够从最早的阶段直接追踪胶质母细胞瘤的进化。出乎意料的是,作者观察到克隆消亡事件的高发生率和克隆大小的渐进性分化,甚至在恶性表型获得后也是如此。计算模拟表明这些动力学是基于克隆的细胞-细胞竞争的结果。通过批量和单细胞转录组分析,结合谱系追踪,作者揭示了Myc转录靶标与克隆大小不平衡的相关性最强。此外,作者还证明了下调Myc的表达足以驱动颅内移植胶质瘤中的竞争动力学。作者的研究结果提供了使用传统回顾性方法无法获得的胶质母细胞瘤进化的见解,突出了在该领域结合克隆追踪和转录组学分析的潜力。本研究于2023年8月于《Cancer Cell》上发表,IF=50.3。
图形摘要

技术路线

主要研究内容
1、胶质瘤诱导barcode library的验证
为在体内诱导肿瘤启动的基因组病变,同时对每个转导的细胞进行特异标记,作者生成了一个致癌逆转录病毒载体的barcoded library。文库中的每个载体包含:(i)一个独特的genetic barcode,(ii)作为报告基因的GFP,(iii)PDGFB致癌基因(图1)。作者将barcode设计为BH模式(一个non-A核苷酸后面接一个non-G核苷酸)的11次重复,最大理论复杂度为322(每个位置上可能的核苷酸数目上升到该位置上的数目),对应约3×1010形式。通过在低感染复数(0.01 MOI)下转导5个独立的NIH3T3细胞培养来测试所获得的逆转录病毒文库的有效复杂性和barcode分布的均匀性。随后通过靶向NGS获得的barcode组成分析显示,其分布没有重大偏差(图1B,C)。库(即,库中功能向量中不同barcode的数量)的有效复杂度为(3±0.15)×104。同时,作者通过处理包含已知比例的两个不同barcode克隆的细胞的不同样本,验证了作者的barcode检索、测序和定量的方法。这项分析表明,即使在代表总样本的10-4倍时,barcode也可以被一致检索和量化。
研究将逆转录病毒文库注射到E14小鼠胚胎的端脑囊泡中。正如已经报道的那样,29个PDGFB的过表达在成年期早期总是导致胶质瘤的发展(图1D)。随着时间的推移,GFP表达标记的胶质瘤的总体质量(即GFP阳性细胞总数)增加(图1E)。在早期出现症状的小鼠中,胶质瘤表现为小而弥漫的多灶性肿块,类似于低级别肿瘤,而在后期出现症状的小鼠中,肿块表现为更大和更密集的(图1F-J)。这些数据与作者先前的观察结果一致,即随着时间的推移,胶质瘤从低级别进展到高级别,并伴随着恶性程度的增加。
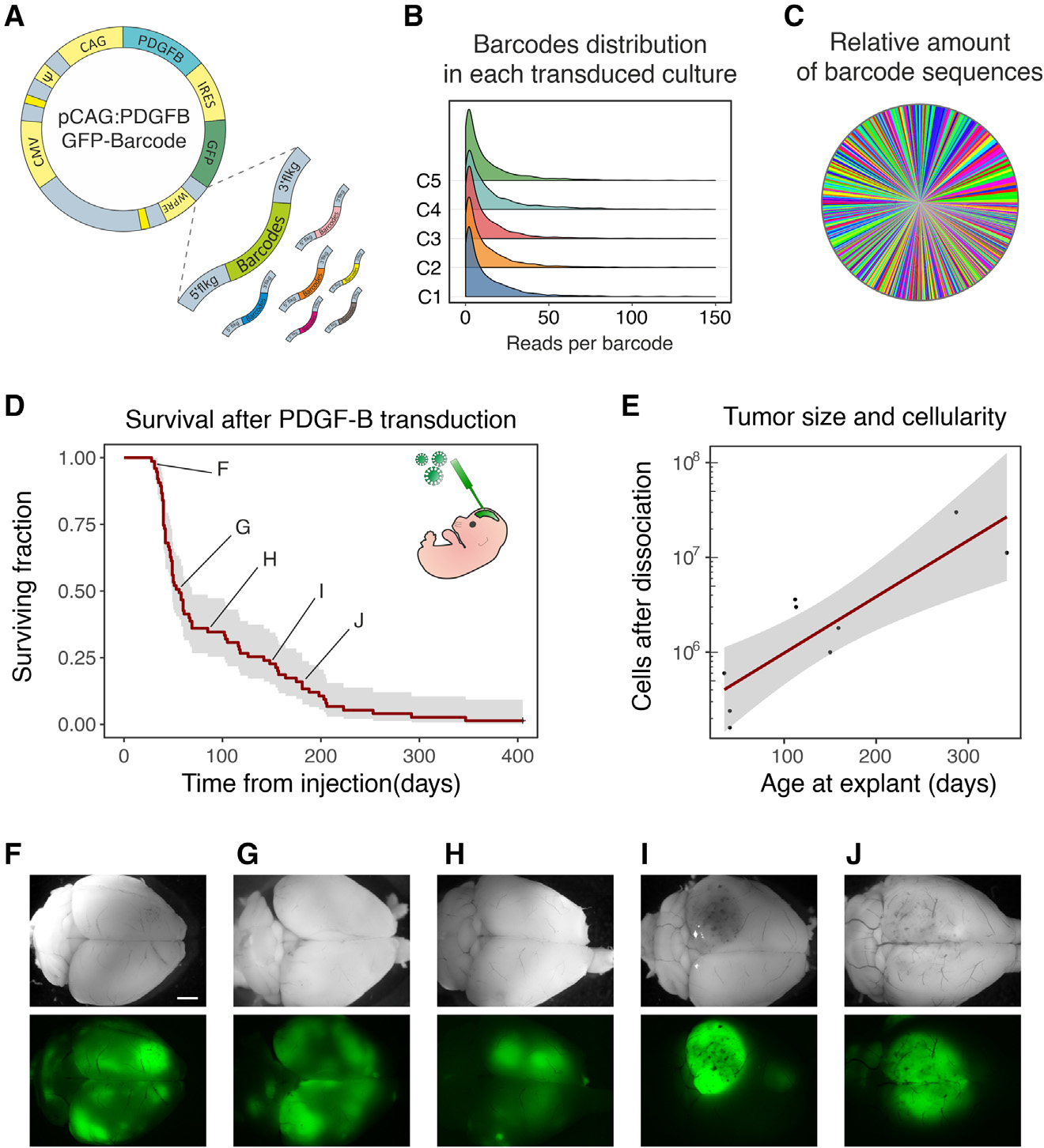
图1 barcode矢量的构建和验证
2、PDGFB诱导胶质瘤的克隆动力学
在注射(n=7)后第4天,从胚胎中提取DNA来估计逆转录病毒转导细胞的数量。每个胚的barcode编号为(16±4)x 103。重要的是,在每个胚胎中,单个barcode的相对丰度是均一的,没有过度代表的barcode,Gini不平等指数为0.51±0.02。该值与原始质粒文库(0.57 ± 0.01)非常接近。随后,对38只小鼠(16窝)在不同时间表现出神经症状的肿瘤块提取DNA进行相同的分析。该分析显示,发育过程中,在出生后151至342天的3只小鼠的主要克隆的数量急剧减少,从33日龄小鼠中的最多495个克隆到单个克隆(图2A,C)。克隆数目的减少一方面与肿瘤质量和细胞数量的增加相呼应(图1E-J),另一方面与它们相对分布的不平衡相呼应,如Shannon-Wiener熵指数(简称Shannon熵)所示,该指数随时间呈指数下降趋向于0(图2B)。这幅图表明,随着时间的推移,以牺牲其他克隆为代价而逐步占优势的原始克隆的子集的选择性扩张。
对这一现象的一个可能的解释是少数细胞成功地克服了biological gate的发生,表现为在向完全恶性肿瘤进展过程中的克隆多样性瓶颈。基于这一假设,晚期发病的胶质瘤,很可能已经通过了这一biological gate,如果移植到成年小鼠的大脑中,不应该面临强大的克隆瓶颈。作者通过将5个迟发性胶质瘤(> 150天)的急性分离细胞移植到同系小鼠(每只胶质瘤至少2只小鼠)中,并比较原发性和继发性胶质瘤的克隆组成,验证了这一假设。移植动物在27 ~ 45天之间出现继发性胶质瘤。随后的克隆分析显示,其中2例移植的胶质瘤是单克隆性的,因此,它们的继发性肿瘤也是单克隆性的。另外3例移植瘤分别由2、3和5个克隆组成,但值得注意的是,每个克隆均产生单克隆继发性肿瘤(图2D)。尽管每个克隆的移植细胞数量(只有一个例外)远高于产生继发性肿瘤所需的最小数量,但这种完全的纯化还是发生了(图2E)。这表明,尽管有前期的进展,肿瘤细胞仍处于强烈的选择性压力之下。然而,这一结果可能受到亲本无性系相对大小的强烈不平等的影响。为了解这种可能的限制,更好地理解在完全进展的肿瘤中正在进行的选择,作者决定用只携带GFP和genetic barcodes的逆转录病毒库标记完全进展的胶质瘤细胞后进行类似的移植实验。作者使用了PDGFB诱导的胶质瘤(CSC-204D39)的原代培养,已知当原位移植到同系小鼠(穿透率> 97 %,n = 64)时,通常会产生继发性肿瘤。将文库转染的细胞移植到成年小鼠(n = 4)的大脑中,当小鼠出现神经症状时收集二次肿瘤。对移植前的细胞和来自二次肿瘤块的细胞进行barcode测序。有趣的是,肿瘤的二次生长伴随着barcode数量的急剧减少,barcode数量从(1.19 ± 0.05)× 104减少到1(n = 4,图2D),这表明即使在进展的胶质瘤中也发生了类似于在胶质瘤进展过程中观察到的瓶颈。这一结果强烈表明,至少一些负责克隆减少的机制可能在获得完全恶性肿瘤后仍在运作,指出存在进一步的肿瘤内选择来源。
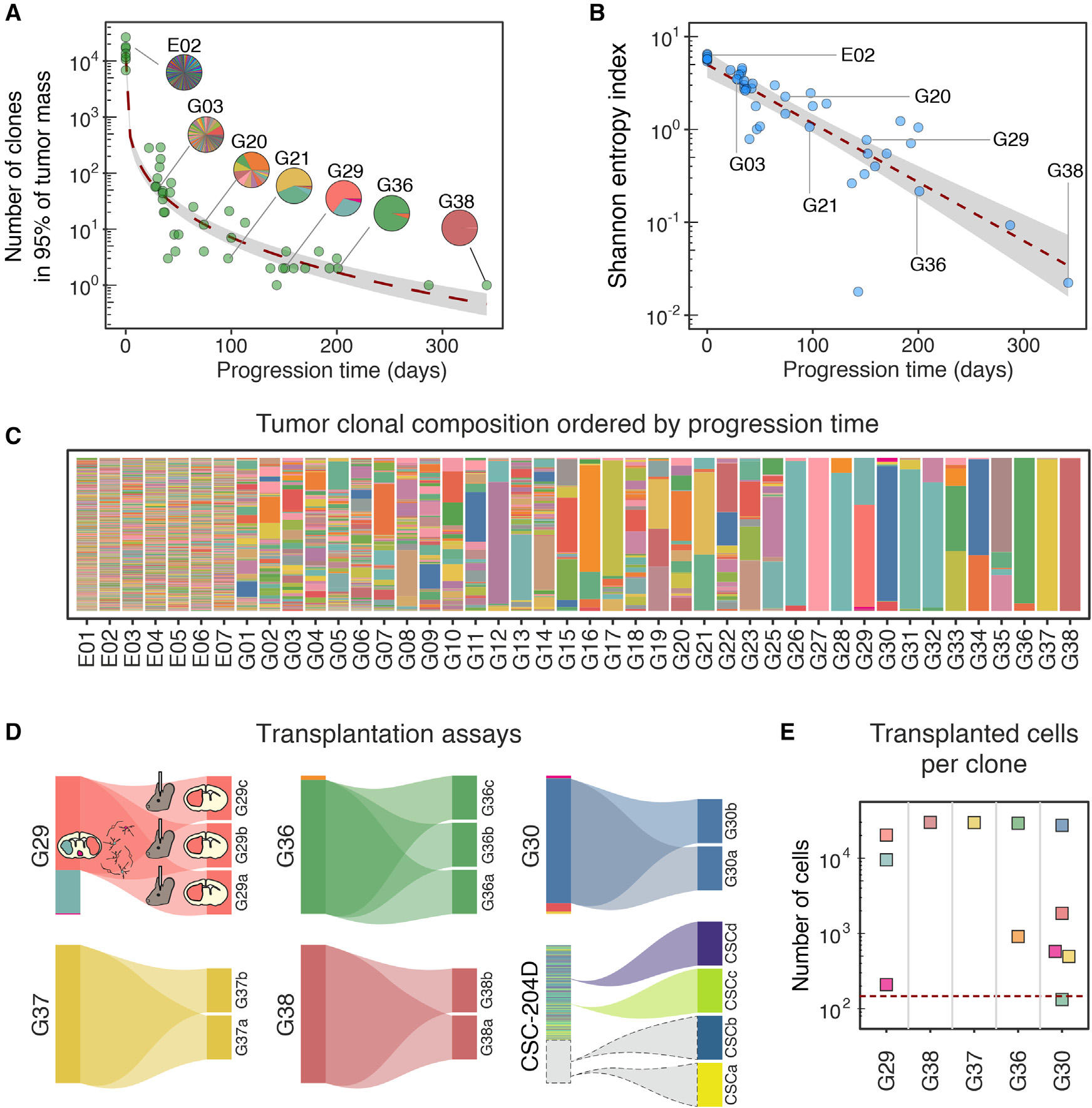
图2 原发性和继发性胶质瘤的克隆组成动态
3、胶质瘤如何失去克隆异质性?
在作者的胶质瘤发生模型中观察到的克隆减少,理论上可以归因于几种不同的非互斥机制:(i)高比率细胞死亡(或细胞周期撤退)导致的统计漂移。细胞死亡或静息可以随机驱动克隆不平衡,从而降低克隆异质性。(ii)细胞周期长度的克隆差异。如果克隆显示不同的细胞周期长度,较快克隆的大小将以指数方式偏离较慢克隆的大小,它们最终将在肿瘤团块中占优势,使其他克隆过度生长,并减少明显的异质性。(iii)基于克隆身份的细胞-细胞竞争。在器官发育和肿瘤发生过程中,特定基因产物表达或活性的遗传性(即,克隆性)差异会引发相邻细胞之间的竞争,为克隆选择提供基础。为研究这些机制对克隆多样性减少的可能影响,作者进行了Monte Carlo simulations,其中相关参数可以单独操纵,以探索它们在塑造克隆动态中的作用。在作者的模拟中,一个“virtual tumor cells”的起始集在一个简化的胶质瘤生长模型上,该模型是由在体外或体内进行的实验措施所产生的现实值。作者开始使用这个框架来检验细胞死亡对克隆多样性降低的影响。首先,作者考虑了一个极限场景,其中所有细胞的周期设置为体外测得的平均长度(22 h),并且在整个模拟过程中细胞死亡概率(pCD)是恒定的。这里作者注意到,当pCD达到一定值时,细胞死亡克服增殖,模拟种群数量减少,最终消失。超过180天,pCD的最高值仍然与肿瘤生长(即每小时约有3 %的死亡细胞)相容,导致克隆多样性从10000个下降到580个克隆(图3A)。与体内(大约4个数量级)相比,这种降低小于1.5个数量级,几乎可以忽略不计。此外,这种细胞死亡率远远高于TUNEL分析所观察到的结果,在体内中,死亡细胞的比例不到0.01 %(图3B)。作者随后引入了一个不同的细胞死亡模型来考虑肿瘤负荷的影响。因此,细胞死亡的概率被建模为肿瘤细胞总数的逻辑函数。作者通过探索nCell50参数来研究不同肿瘤负荷强度的影响。该参数对应于负责一半肿瘤负荷效应的细胞数(即Logistic曲线的拐点)。即使在这种情况下,细胞死亡要么太明显,不允许肿瘤生长,要么太轻微,不能重复实验观察到的克隆多样性减少的程度(图3C)。同样,在这种情况下,可检测到的克隆减少(每小时约有3.2%的死亡细胞)所对应的细胞死亡净百分比与作者的TUNEL分析结果不一致。上述结果排除了在胶质瘤发生过程中高细胞死亡率可以自行解释观察到的克隆动态的可能性。
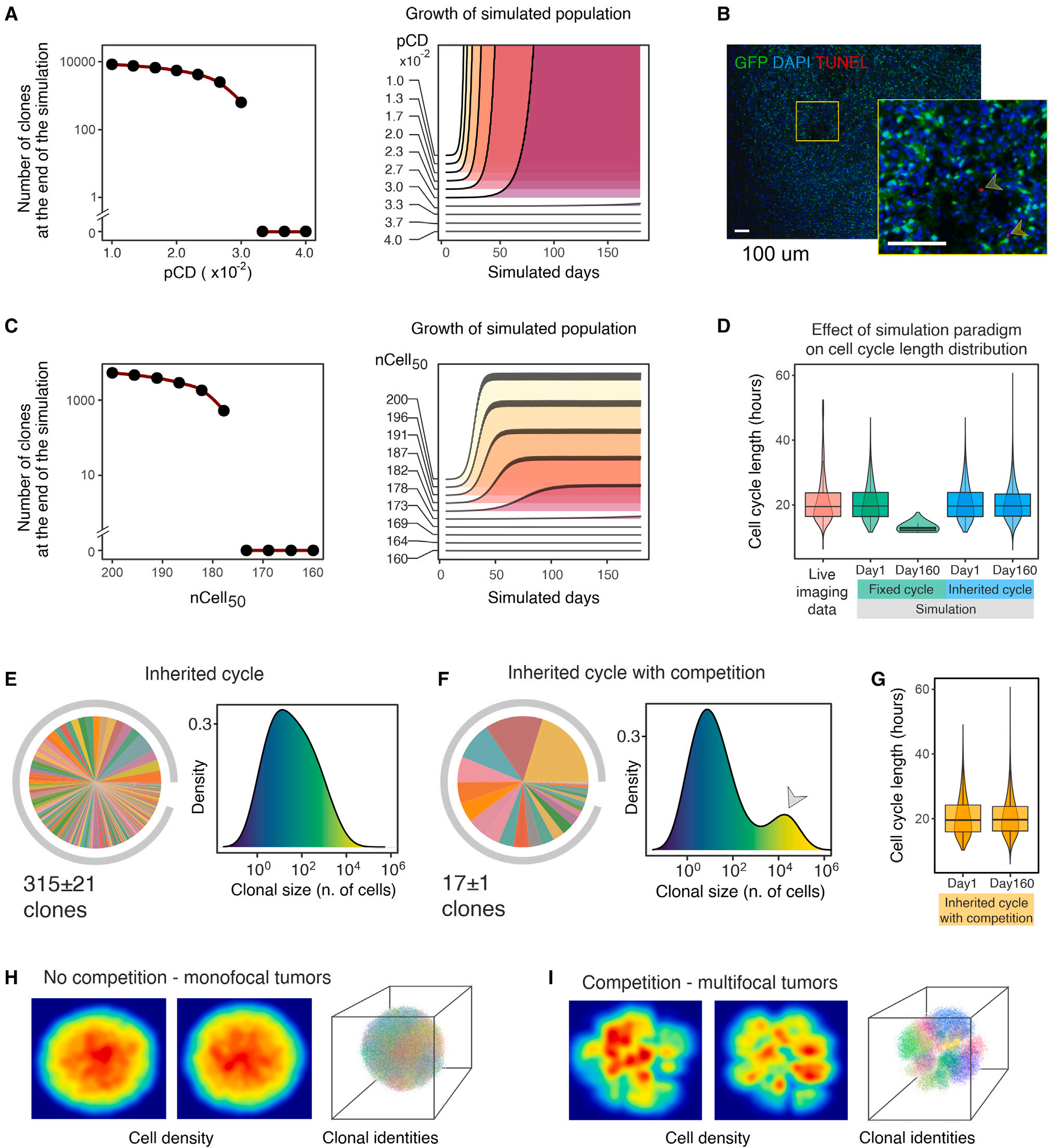
图3 胶质瘤生长的计算分析
4、局部细胞密度和细胞周期长度波动
接下来,作者增加了模型的复杂性,包括细胞周期长度的克隆差异。作者从理论上的极端假设出发,即每个子细胞从其母细胞继承一个固定的、未被修饰的细胞周期长度。在此假设下,通过初始化由视频延时显微镜测量的细胞周期长度分布的原始群体,模拟结果产生了与作者在体实验数据不兼容的图片。更快的循环细胞的选择性扩张导致了中位细胞周期长度随时间的下降,下降到约14 h,这比实验观察到的明显缩短(图3D)。尽管这种不切实际的行为,但在160个模拟日中,克隆的数量从10,000个变化到约200,比作者所观察到的减少得更少。更真实的模拟包括细胞分裂后细胞周期长度的变化。在这种范例中,细胞继承其父代的细胞周期长度,加上一个正态分布的随机值。这种模拟结果使细胞周期长度的分布更接近实验数据(图3D)。在此条件下,同样地,虽然几个生物学参数正确地再现了(如总体肿瘤生长率,细胞周期长度和肿瘤体积的分布),但在160个模拟日中,克隆数量从10000个下降到约665个(图3E)。再次,该结果与实验数据不符。因此,克隆多样性降低似乎不太可能仅仅是由于细胞周期长度差异造成的。
5、细胞与细胞之间的竞争
最后,作者在上述模型中加入了基于克隆身份的细胞-细胞竞争的贡献。通过区分同胞(同一克隆)和无关(不同的克隆)细胞,每个细胞的适合度受到其邻近细胞的影响,这取决于它们各自的克隆身份。特别地,作者建模了一个场景,对于任意给定的细胞,不相关的邻居比兄弟姐妹对细胞死亡概率的影响更大。在这个模拟中,作者获得了从10000到108的减少,值得注意的是,其中只有17 ± 1占总人口的95 %,Shannon熵为2.5 ± 0.1。此外,由于超级竞争者克隆的出现,克隆大小呈现多峰分布(图3F)。值得注意的是,触发局部克隆竞争并不影响细胞周期长度分布(图3G),并导致模拟的肿瘤肿块具有与实际体内肿瘤大体相似的大小和拓扑结构,显示出密集的分叶被几乎无瘤的区域(图3H,I)隔开。没有其他经过检验的假设导致如此明显的克隆不平衡和多样性减少。
6、进展中的肿瘤细胞动力学(如何竞争)
为深入了解细胞-细胞竞争的可能机制,作者对PDGFB诱导的不同时间收获的胶质瘤进行了RNA测序(RNA sequencing, RNA-seq)分析,(n = 4只小鼠,出生后39天)之前或神经症状发作时(n = 7只小鼠,出生后40 ~ 176天)。由于该barcode序列在结构上与高表达的PDGFB-IRES-GFP转录本相连,因此可以通过RNA-seq分析高效检索。这使得作者可以直接估计每个肿瘤的Shannon熵,并将其与性状相关基因(trait-associated gene,TAG)分析中的基因表达相关联。然后使用TAG衍生的排序来驱动基因集富集分析(GSEA),在Hallmark基因集上进行,克隆复杂性的降低与Myc信号直接相关(Myc Targets V1和E2F靶点紧随Myc靶点V2,p < 10-5,图4B,C)。这个结果是非常有意义的,因为Myc信号传导是涉及细胞-细胞竞争的少数已描述的机制之一。Myc表达本身是超级竞争者克隆的特征,与Shannon熵显著负相关,这表明Myc作为克隆丢失(Pearson相关性: 0.85,p < 0.001,图4D)的驱动因子的潜在作用。作者推测,Myc水平的升高可能是由于对表达较高水平Myc的细胞进行选择,从而逐渐富集的结果。然而,考虑到在完全进展的肿瘤中也有在单个克隆中持续选择的迹象(图2D),作者想知道即使完全纯化,单克隆肿瘤是否会在细胞水平上保持异质性的Myc表达。为解决这个问题,作者分离了两个单克隆肿瘤,并通过基线稀释克隆分离亚克隆培养物。然后,作者量化了每个肿瘤10个亚克隆的Myc表达,并将差异与亲代培养中观察到的波动进行了比较。在这两种肿瘤中,作者观察到亚克隆培养物中Myc表达的异质性高于亲代肿瘤(图4E)。作者利用RNA-seq数据佐证了这一证据,以提示在胶质瘤进展过程中克隆内异质性的维持。在胶质瘤进展的早期阶段(PDGFB转导后小于45天)中,大多数可检测到的突变并不是在所有的胶质瘤细胞(等位基因频率中位数= 0.20)中共享的。在后期,出现了一组广泛的突变(等位基因频率中位数= 0.93),可能是某些克隆扩张的迹象,与作者之前的数据一致。尽管如此,低频突变仍然是绝大多数,这表明基因组变异在扩大的克隆中不断涌现(图4F)。
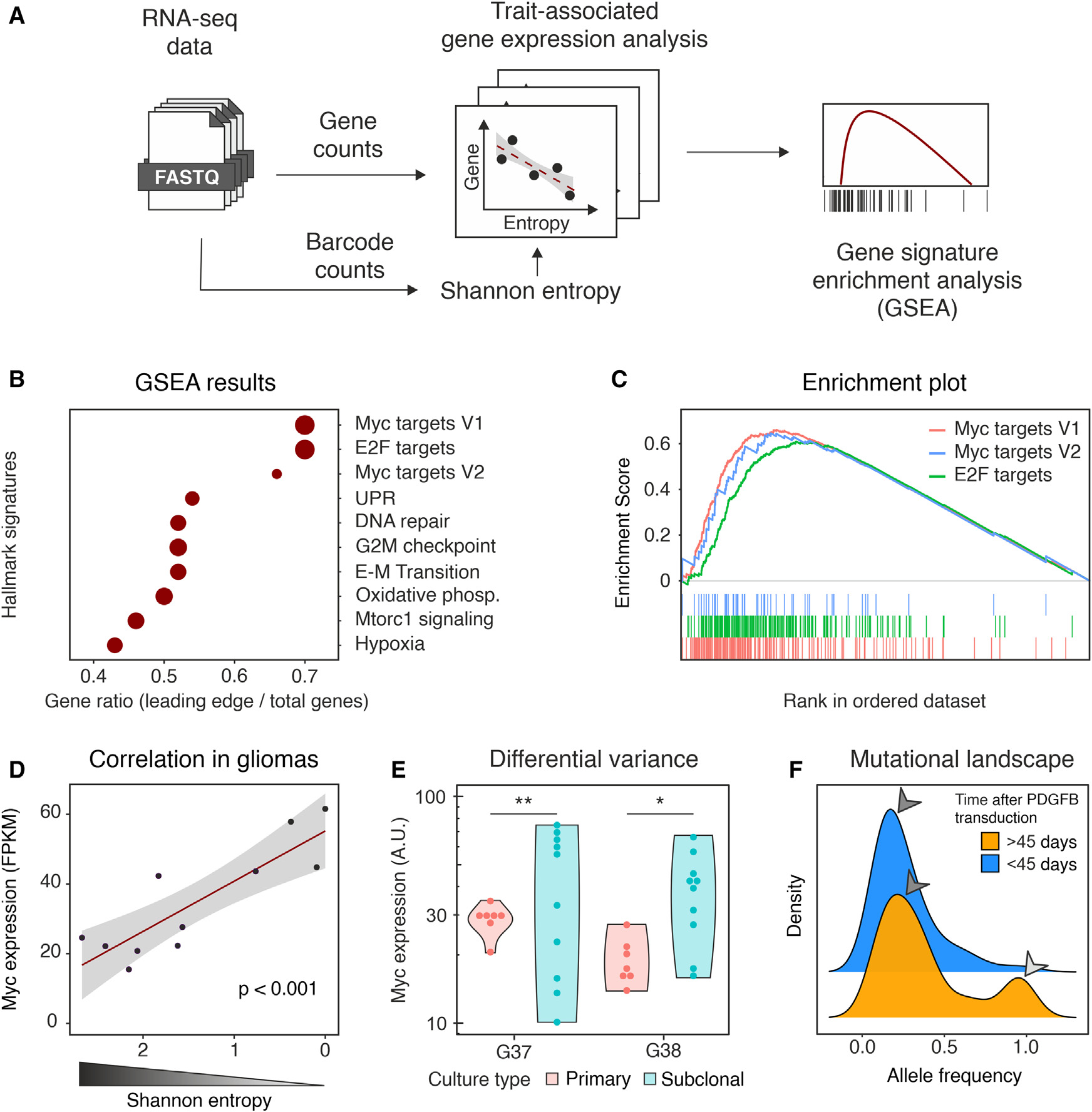
图4 与进展性胶质瘤bulk RNA-seq相关的克隆分析
7、胶质瘤的早期克隆分化
为对克隆间竞争的潜在驱动因素进行更高分辨率的搜索,作者对胶质瘤进行了单细胞RNA-seq(sc RNA-seq)分析,重点关注可能存在许多不同克隆且其大小开始发散的阶段。根据作者之前的分析,这对应于出生后大约40天(图2A)。分离3个独立的PDGFB诱导的胶质瘤细胞,通过荧光激活细胞分选分离出GFP阳性的细胞组分,并用Chromium 10X进行单细胞分选。同时,通过对barcode区域的靶向DNA测序,分析了分离细胞的代表性部分(5 %)。后一种分析使作者能够确定每个barcode克隆的大小。然后,在scRNA-seq数据中搜索本分析中发现的barcode,将细胞的转录组图谱与克隆信息联系起来,并将其与属于的克隆的大小和身份联系起来(图5A-5C)。数据显示,克隆通常由属于不同转录簇的细胞组成(图5D)。值得注意的是,在GSEA分析中,当比较属于小克隆(单个细胞占肿瘤质量的30%以下,累计约占测序细胞总数的二分之一)的细胞和属于大克隆(分别代表至少30 %的肿瘤块,并累积组成另一半测序细胞)的细胞时,最富集的基因签名是Myc信号和相关的E2F-靶标,再次是(p < 10-4,见图5E、F)。这增加了Myc-E2F轴在控制克隆大小方面的作用的证据,并指出其在胶质瘤发生过程中塑造细胞-细胞竞争动力学的重要性。进一步分析显示,不同大小克隆的细胞中转导的PDGFB转基因表达没有差异,而大克隆的细胞中Myc表达有较高的趋势(图5G)。
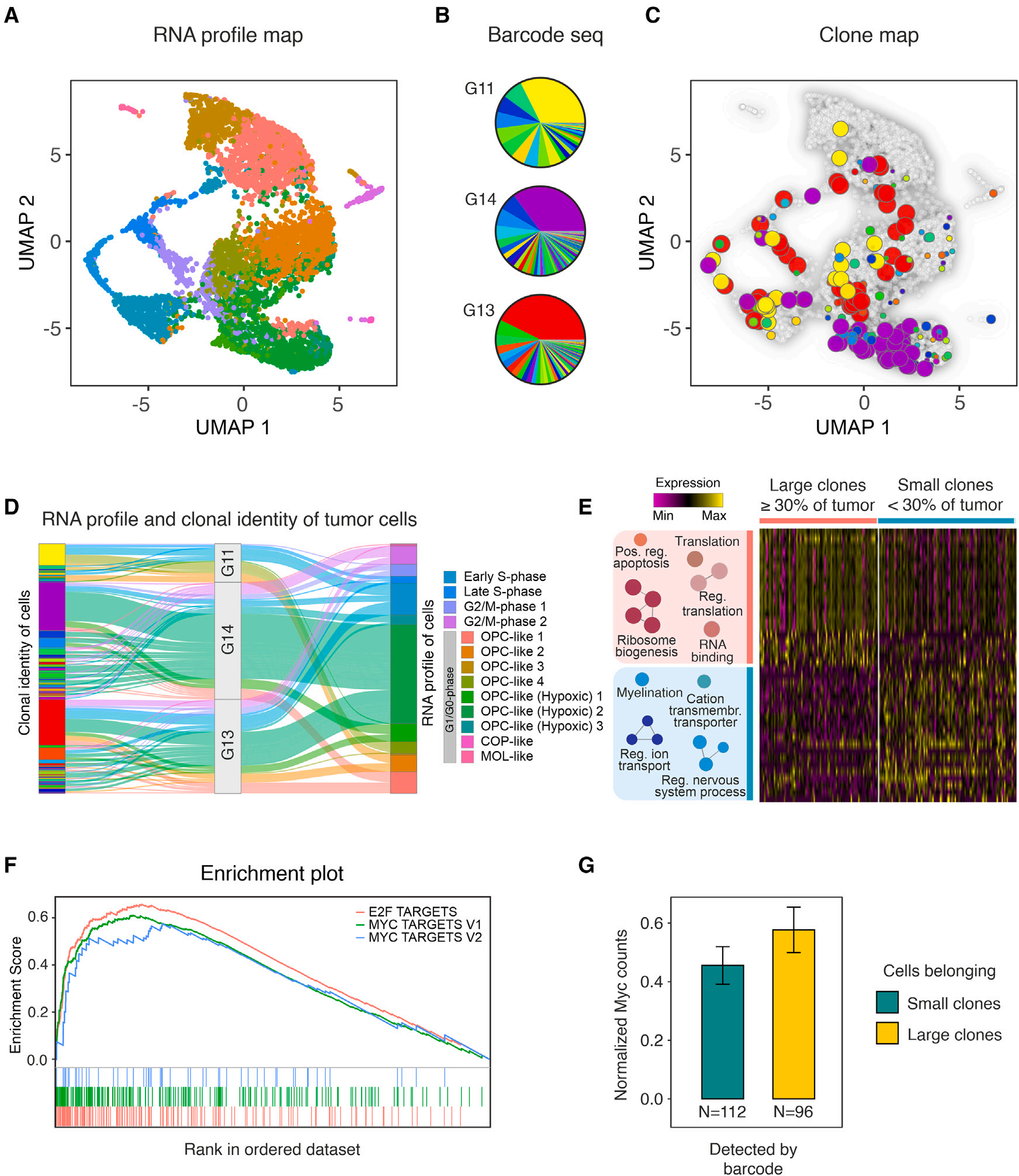
图5 早期胶质瘤的单细胞RNA-seq分析
8、胶质瘤细胞中Myc介导的竞争
然后,作者从功能上评估了Myc表达水平在基于克隆的竞争中的作用。作者使用CSC-204D细胞,即PDGFB驱动的胶质瘤原代培养细胞,用于图2D所示的实验。作者通过稳定转导两种不同的携带GFP的逆转录病毒载体和一种旨在抑制Myc表达的人工miRNA(amiRNA)来构建CSC-204D细胞。作者通过qPCR验证了两种不同的anti-Myc amiRNAs(miRMyc16和miRMyc19)介导的敲低,分别降低到原来的52 ± 4%和61 ± 6%(图6A)。这种调节水平与作者在原代胶质瘤细胞中观察到的情况非常相似(图4D)。随后,作者通过MTT实验比较了Myc敲低(Myc-knockdown,Myc-KD)细胞和转染scramble miRNA的细胞的体外生长速率,培养物的复制时间无明显变化(图6B,C)。作者进行了移植实验以测试Myc-KD胶质瘤细胞单独移植或与Myc野生型(Myc-wt)细胞混合移植时的行为。特别地,作者设计了两个互补的实验来推断Myc在胶质瘤生长和克隆动力学中的作用:在第一个实验中(实验1),作者将Myc-KD细胞(用RFP和GFP标记)单独原位移植到小鼠大脑中,并检查它们是否可以作为它们的亲本群体产生继发性胶质瘤。在第二个实验(实验2)中,作者原位移植相同数量的Myc-KD细胞与等数量的Myc-wt细胞(仅用RFP标记)混合,检测这两个亚群是否可以共同生长,或者Myc-KD细胞在继发性胶质瘤中是否会被淘汰。作者推断,实验1中胶质瘤的发展,以及实验2中Myc-wt细胞的纯化趋势,表明Myc在胶质瘤进展过程中发挥了驱动竞争的作用(图6D)。事实上,实验1中所有移植的小鼠都发生了继发性胶质瘤(7/7)。值得注意的是,胶质瘤的发生时间与实验2相同,Myc-KD和Myc-wt胶质瘤细胞在体内的成瘤能力和生长速度没有差异(图6E-6G和6J)。在实验2中,所有的移植小鼠(7/7)都发生了继发性胶质瘤,并且Myc-wt细胞被高度纯化。几乎没有Myc-KD细胞(RFP和GFP双标记)被发现(图6H-K)。在作者使用相同细胞进行的体外实验中也得到了类似的结果。作者将Myc-KD(或scrambled miRNA control)细胞和Myc-wt细胞以1:1的比例混合在U型多孔板中形成球体。然后作者在三个时间点监测他们的生长情况。
Myc-KD细胞被Myc-wt细胞取代的比率显著高于对照胶质瘤细胞(图6L-N)。总之,这些结果证实了作者提出的Myc在胶质瘤发生过程中驱动基于克隆的竞争的作用。
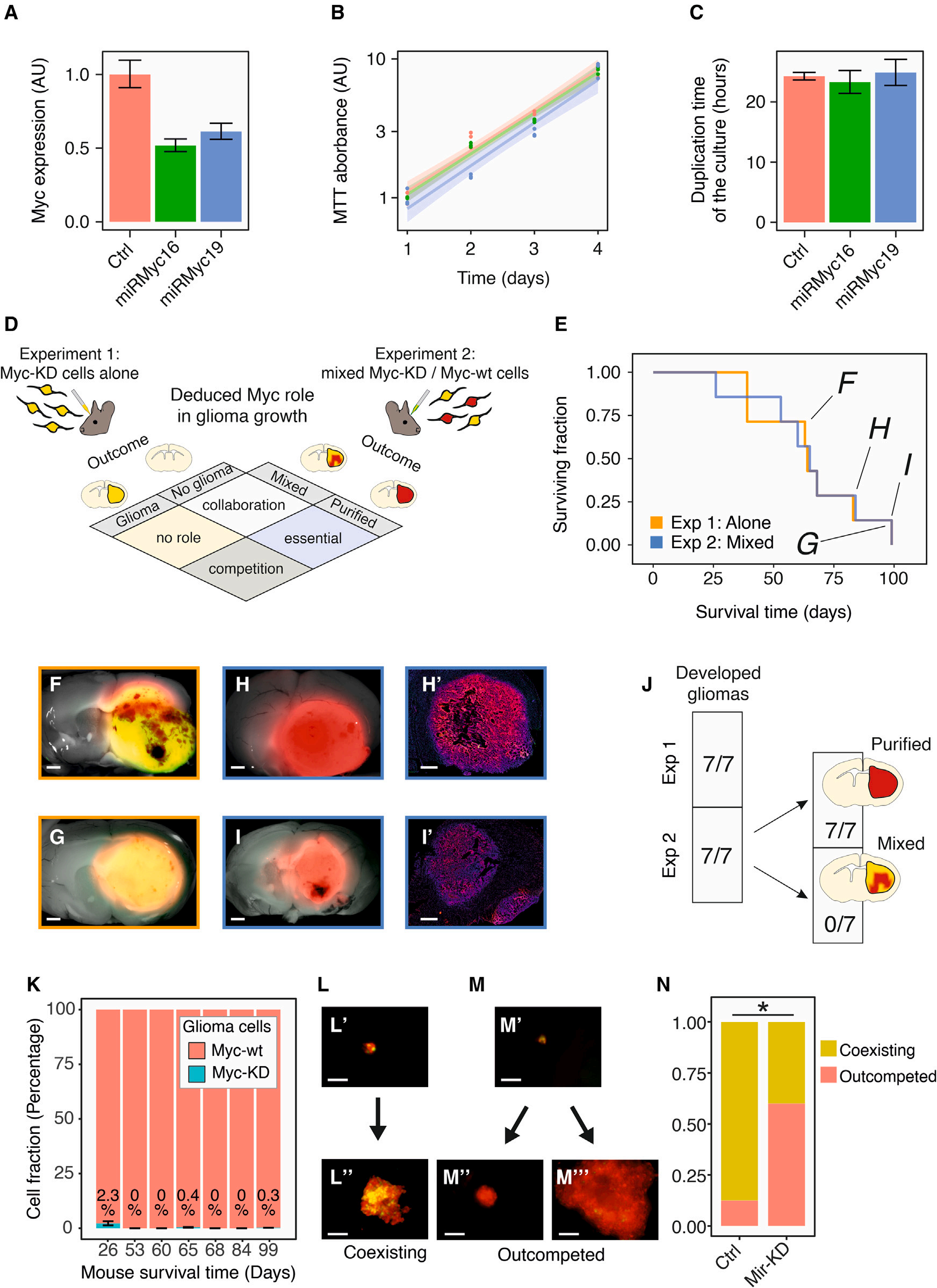
图6 Myc modulation驱动胶质瘤生长的竞争动态
实验方法
C57BL/6J小鼠,胶质瘤细胞原代培养,胶质瘤细胞的极限稀释克隆,胶质瘤细胞中Myc下调,免疫染色和TUNEL试验,定量PCR,逆转录病毒barcoding libraries production,Barcode sequencing,Bulk RNA sequencing,scRNA-seq,基因表达分析,基因组变异分析
参考文献
Ceresa D, Alessandrini F, Lucchini S, Marubbi D, Piaggio F, Mena Vera JM, Ceccherini I, Reverberi D, Appolloni I, Malatesta P. Early clonal extinction in glioblastoma progression revealed by genetic barcoding. Cancer Cell. 2023 Aug 14;41(8):1466-1479.e9. doi: 10.1016/j.ccell.2023.07.001. Epub 2023 Aug 3. PMID: 37541243.