






LncRNA NORAD/miR-224-3p/MTDH轴通过促进β-catenin核积累,参与食管鳞状细胞癌CDDP耐药
食管鳞状细胞癌(ESCC)是我国食管癌的主要亚型,占食管癌患者的90%以上。目前,顺铂(cis-diamminedichloro-platinum,CDDP)是治疗ESCC的首选药物。CDDP耐药是一种恶性的生物学特性,其调控机制复杂。长链非编码RNA (lncRNAs)的异常调节是CDDP耐药的关键调控因子。然而,本研究探讨ESCC中参与CDDP耐药的LncRNA并阐明其临床意义和功能机制。本文于2021.12月发表于《Molecular Cancer》, IF=15.302。
技术路线:

主要结果
1.在ESCC中,NORAD高表达与CDDP耐药相关
基于GEO数据库,作者首先分析了CDDP敏感和耐药的食管癌组织中差异表达的基因,并筛选5个在CDDP耐药的组织中差异表达的LncRNA(Fig. 1a)。这5个LncRNA在CDDP耐药的ESCC组织中NORAD表达量明显高于CDDP敏感的ESCC组织(Fig. 1b)。 TCGA数据库分析发现NORAD在ESCC组织中的表达高于正常组织,提示NORAD可能促进食管鳞癌的发生ESCC进展(Fig. 1c)。荧光原位杂交显示在CDDP耐药的ESCC组织中,NORAD的表达量明显高于CDDP敏感的ESCC组织(Fig. 1d)。 临床分析发现,NORAD表达量越高患者预后越差(Fig. 1e)。ROC曲线显示NORAD可能是ESCC患者CDDP耐药的预测标志物(Fig. 1f)。风险模型显示发现NORAD表达和肿瘤分期都是ESCC患者无病生存期和总生存期的独立危险因素(Fig. 1g)。
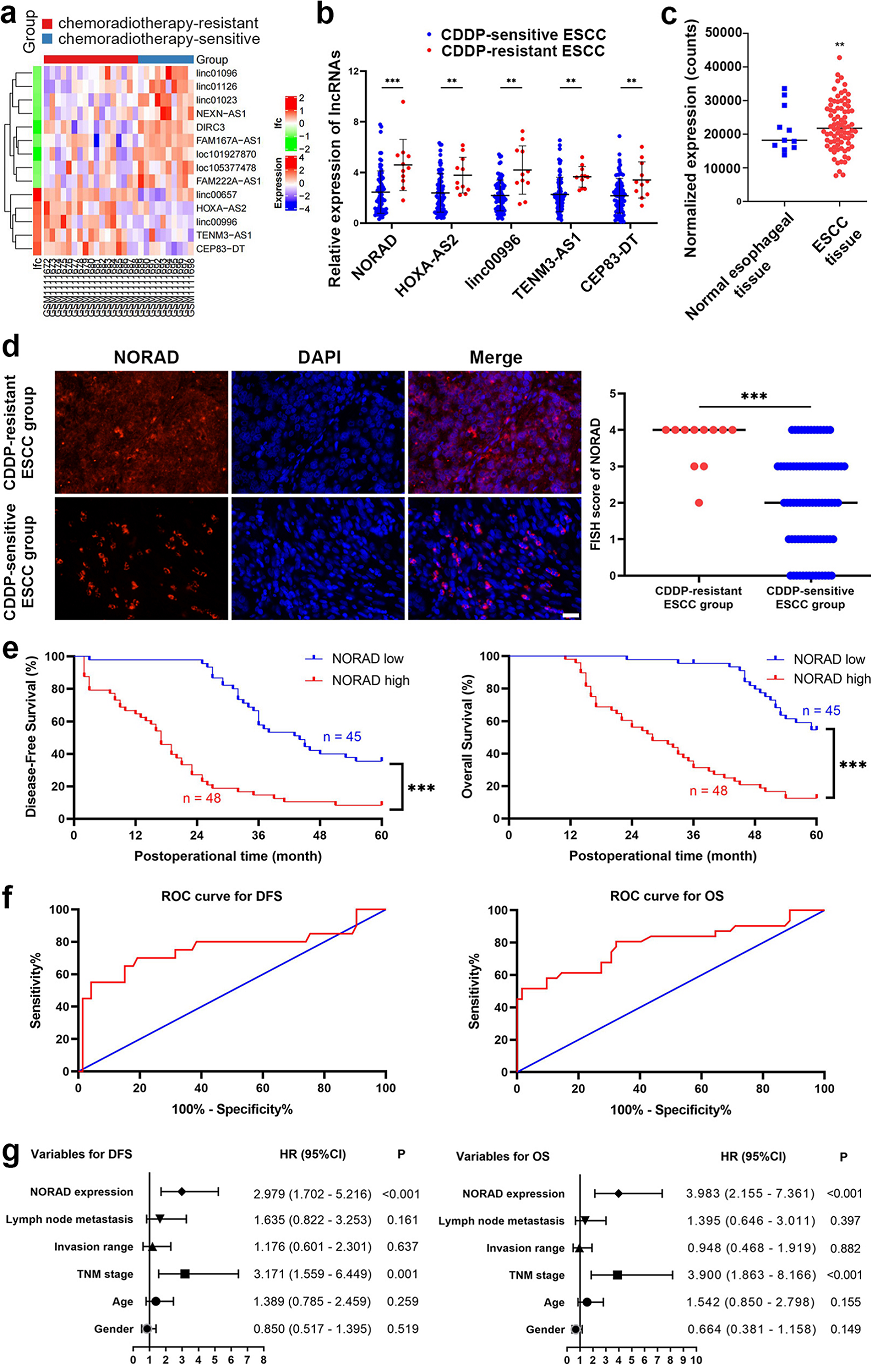
Fig 1 在CDDP耐药的ESCC组织中,NORAD表达增加,与患者预后不良相关
2.NORAD促进体外ESCC CDDP耐药
为了进一步研究CDDP耐药在ESCC中的作用机制,作者采用CDDP敏感的亲本ESCC细胞系(KYSE30和TE1)及其等基因耐药CDDP细胞 (KYSE30/CDDP-R和TE1/CDDP-R)用于后续研究。结果发现KYSE30/CDDP-R和TE1/CDDP-R中,NORAD显著高于相应的亲本细胞(Fig. 2a)。通过荧光原位杂交检测到NORAD主要位于ESCC细胞的细胞质中(Fig. 2b)。IC50是指在用药后存活的细胞数量减少一半时所需的药物浓度,在KYSE30/CDDP-R和TE1/CDDP-R细胞系中敲低NORAD,在KYSE30和TE1细胞系中过表达NORAD,结果表明,下调NORAD表达显著降低了KYSE30/CDDP-R和TE1/CDDP-R中的IC50值,表明NORAD降低了耐药细胞的敏感性(Fig. 2c)。此外,过表达NORAD显著降低了KYSE30和TE1细胞对CDDP的敏感性(Fig. 2d)。此外,NORAD在耐药细胞中敲低之后抑制了集群形成能力,表明NORAD基因敲除部分恢复了这些细胞对CDDP的敏感性(Fig. 2e);与空载体(EV)组相比,用CDDP处理TE1细胞时,NORAD过表达对TE1细胞的增强作用更为显著(Fig. 2f)。作者接下来了检测细胞凋亡率,以进一步确定NORAD对ESCC细胞CDDP耐药的影响。在CDDP耐药细胞中,抑制NORAD显著增加了CDDP诱导的KYSE30/CDDP-R和TE1/CDDP-R细胞凋亡(Fig. 2g);NORAD过表达显著降低了CDDP诱导了KYSE30和TE1细胞中的细胞凋亡(Fig. 2h)。 在NORAD敲除的耐药细胞中,加剧了KYSE30/CDDP-R和TE1/CDDP-R细胞中CDDP诱导G1期阻滞的情况。当在KYSE30和TE1细胞中过表达NORAD,CDDP诱导G1期阻滞情况被缓解(Fig. 2i)。γH2AX 是DNA损伤标志物,cleaved – caspase-3是凋亡标志物。在KYSE30和TE1细胞中干扰NORAD后,细胞中γH2AX和cleaved - caspase-3的表达显著增加(Fig. 2j);CDDP诱导的DNA损伤标志物γH2AX和凋亡标志物cleaved - caspase-3在KYSE30和TE1细胞中的表达水平也被NORAD过表达降低(Fig. 2k)。
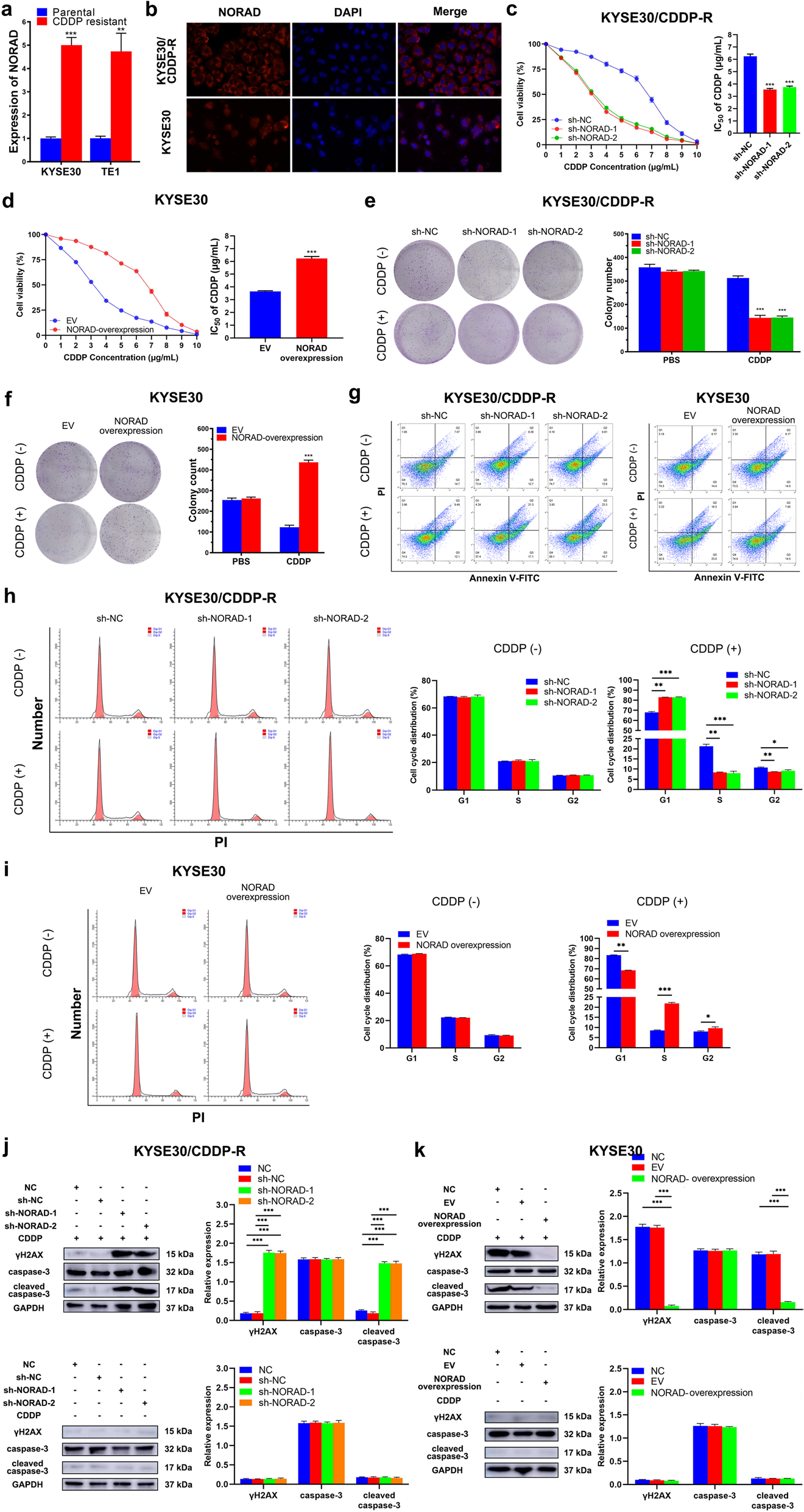
Fig 2 NORAD有助于ESCC的CDDP耐药性
3. NORAD作为miR - 224 - 3p海绵调节ESCC细胞MTDH的表达
为找到参与调控CDDP耐药的miRNA,作者首先通过数据库在CDDP敏感和耐药的组织中找到3个差异表达的miRNAs(Fig. 3a)。其中miR-224-3p可能与NORAD存在结合(Fig. 3b)。在KYSE30/CDDP-R和TE1/CDDP-R细胞中,敲除NORAD之后3个候选差异表达的miRNAs中只有miR-224-3p的表达增加(Fig. 3c)。 在5个片段克隆中敲除NORAD,miR-224-3p在KYSE30/CDDP-R和TE1/CDDP-R细胞中的表达增加均显著上调(Fig. 3d)。表达量分析发现在ESCC组织中,只有miR-224-3p与NORAD呈负相关(Fig. 3e)。此外,KYSE30/CDDP-R和TE1/CDDP-R细胞中的miR-224-3p表达低于相应的亲本细胞(Fig. 3f)。在药物敏感细胞中过表达NORAD,miR-224-3p表达量降低(Fig. 3f)。亚细胞定位检测发现NORAD和miR-224-3p都是定位于ESCC细胞细胞质中(Fig. 3h,i)。将携带NORAD-WT或NORAD突变型的荧光素酶报告基因转染到,KYSE30/CDDP-R和TE1/CDDP-R细胞中,再转染miR-224-3P模拟物。结果表明,在转染NORAD-WT和第二个NORAD突变型中,miR-224-3p模拟物能降低,KYSE30/CDDP-R和TE1/CDDP-R细胞中NORAD的荧光素酶活性。相比之下,转染NORAD- mut #1的KYSE30/CDDP-R和TE1/CDDP-R细胞中,NORAD的荧光素酶活性没有被miR-224-3p模拟物降低(Fig. 3j)。此外,RIP-PCR结果显示NORAD和miR-224-3p在ESCC细胞中形成RNA诱导的沉默复合体(RISC)(Fig. 3j)

Fig 3 NORAD充当海绵吸附ESCC中的miR-224-3p
4. MTDH是miR-224-3p的直接靶点
生信工具预测了miR-224-3p潜在调控的405靶点,通再过生信靶点预测和耐药和敏感的ESCC组织之间的差异mRNA的分析,找到1个重叠区,也就是MTDH(Fig.4ab)。免疫组化检测到MTDH主要位于ESCC细胞的细胞质中(Fig.4c)。接下来检测了CDDP耐药和敏感食管癌细胞中MTDH的表达。发现,KYSE30/CDDP-R和TE1/CDDP-R细胞中MTDH表达明显高于相应的亲本细胞(Fig.4 d),生信预测到MTDH的3’-UTR区中有4个位点有可能与miR-224-3p结合,接下来作者构建了MTDH-3′-UTR的4个不同的突变序列(e.g. MTDH-3′-UTR-Mut#1, MTDH-3′-UTR-Mut#2,MTDH-3′-UTR-Mut#3 and MTDH-3′-UTR-Mut#4) (Fig.4 e)。然后将携带MTDH-3-UTR-WT或MTDH-3-UTR Mut的报告基因导入耐药细胞,再共转染miR-224-3p模拟物(Fig.4 f)。转染miR-224-3模拟物后显著降低了MTDH-3-UTR-WT,以及其中3个MTDH-3’--UTR-mut的耐药细胞中的MTDH的荧光素酶活性。说明miR-224-3p与MTDH-3′-UTR-Mut#1结合。WB得到相同的结果(Fig.4 g)。接下来为了研究NORAD是否通过吸附miR-224-3p上调MTDH的表达,作者在KYSE30/CDDP-R和TE1/CDDP-R细胞中用shRNA敲除NORAD,并转染miR-224-3p抑制剂。结果显示, NORAD基因敲除后MTDH的表达明显降低,而转染miR-224-3p抑制剂后抑制了这种效果(Fig.4 h)。接下来,在KYSE30和TE1细胞中过表达NORAD显著增加了MTDH的表达,而转染miR-224-3p模拟物部分挽救了这种上调(Fig.4 i)。相关性分析发现miR-224-3p与NORAD和MTDH呈负相关, NORAD与MTDH呈正相关(Fig.4i)。
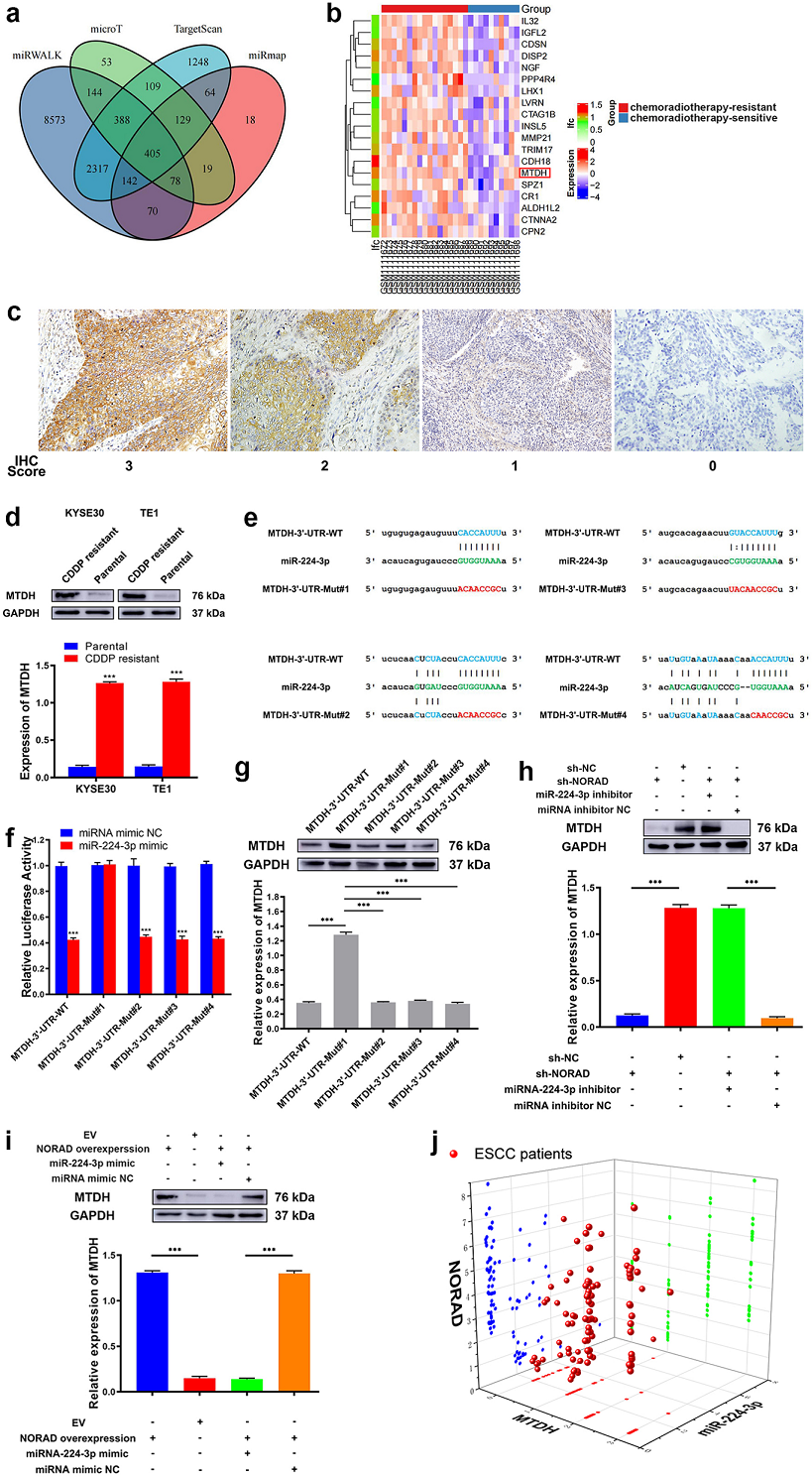
Fig4 MTDH是miR-224-3p的直接靶点
5. NORAD/miR‑224‑3p/MTDH轴通过调节B-活性进而促进耐药以及ESCC进展
接下来,作者用CDDP处理KYSE30/CDDP-R和TE1/CDDP-R细胞,以确定NORAD/ miR-224-3p/MTDH轴在ESCC细胞中的作用。敲低NORAD可部分消除KYSE30/CDDP-R和TE1/CDDP-R细胞中的CDDP耐药性,而miR-224-3p抑制剂则增强了耐药性。随着NORAD过表达,KYSE30和TE1细胞中的敏感性下降,而miR-224-3p模拟物中和了这一现象(Fig 5a)。 此外,随着敲除NORAD,CDDP诱导的KYSE30/CDDP-R和TE1/CDDP-R细胞中的凋亡率增加,这种效果通过共转染miR-224-3p抑制剂来拯救;同时,过表达NORAD,CDDP诱导的TE1和KYSE30细胞凋亡率降低,这种效果能随着转染miR-224-3p模拟物拯救(Fig 5b)。此外,敲低NORAD促进了CDDP诱导的诱导G1期阻滞,而miR-224-3p抑制剂挽救了这种增长;同样,miR-224-3p模拟物部分中和了在KYSE30和TE1细胞中过表达CDDP诱导的G1期阻滞(Fig 5c-d)。为确定NORAD/ miR-224-3p/MTDH轴可能的下游信号通路。于是通过WB检测转染NORAD和miR-224-3p模拟物细胞中不同通路的相关蛋白表达量。发现敲除NORAD下调了磷酸化-β连环蛋白的表达,而且这种下调趋势能被miR-224-3p抑制剂中和(Fig 5e)。接下来分析了NORAD对β连环蛋白在ESCC细胞中的定位情况。WB结果显示敲低NORAD基因后,ESCC细胞的细胞核中β连环蛋白降低,细胞质中β连环蛋白升高(Fig 5f)。此外,作者通过免疫荧光染色观察NORAD/miR-224-3p/MTDH轴对ESCC细胞中β-catenin亚细胞位置的影响。与WB结果一致(Fig 5G)。敲除NORAD或MTDH再共转染miR-224-3p模拟物可减少耐药细胞中β连环蛋白的核积累(Fig 5G)。
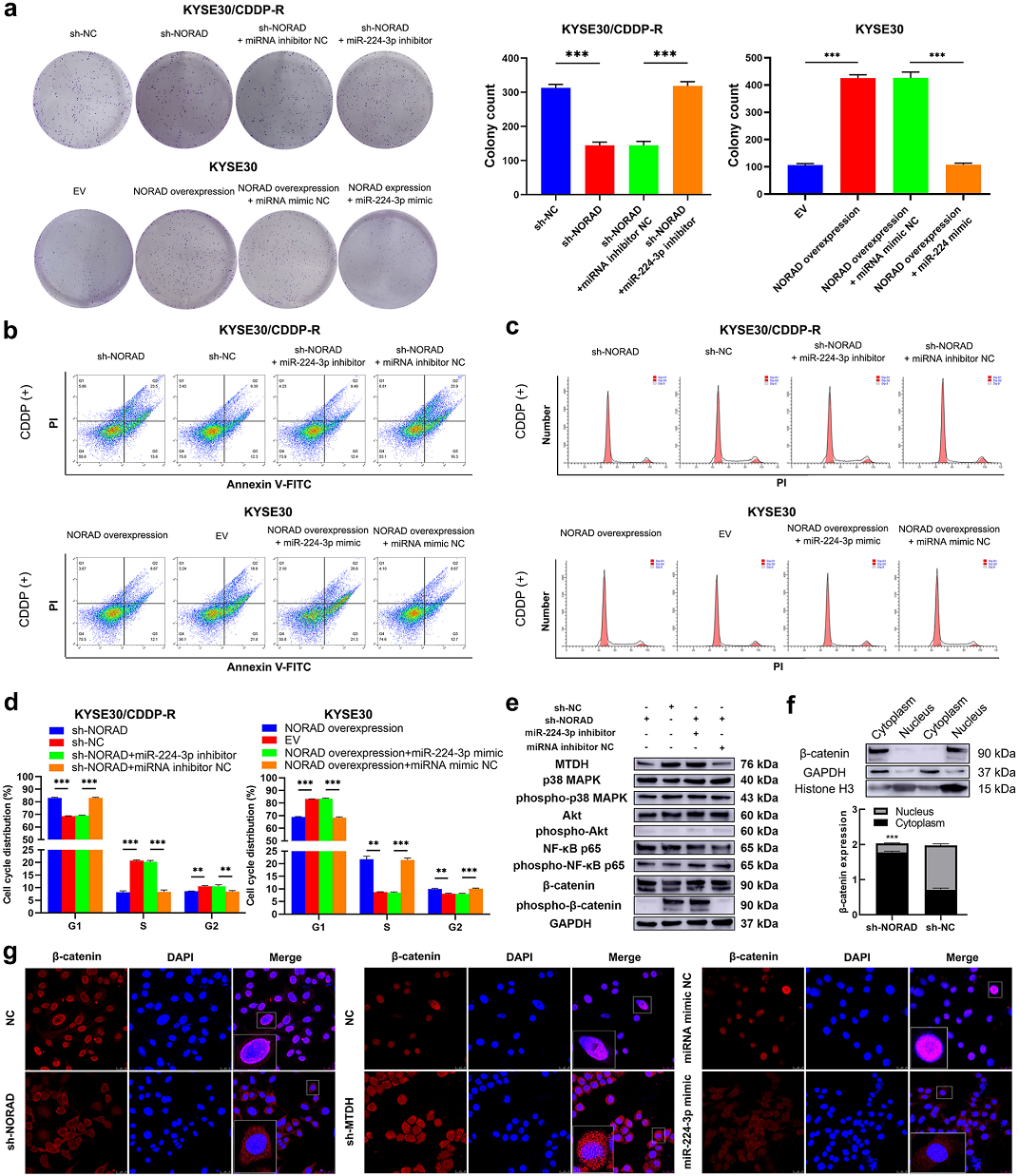
Fig 5 NORAD/miR-224-3p/MTDH轴通过促进β-catenin的核积累来促进ESCC细胞对CDDP的耐药性
6. NORAD/miR-224-3p/MTDH轴促进ESCC细胞的进展
通过伤口愈合试验和Transwell实验发现敲低NORAD抑制了KYSE30/CDDP-R和TE1/CDDP-R细胞的迁移和侵袭,而miR-224-3p抑制剂减弱了这种抑制(Fig 6A,B)。 敲低NORAD后,检测迁移和入侵相关因子的表达量,发现E-cadherin表达量增高,N-cadherin和MMP9表达量降低,而miR-224-3p抑制剂则中和了这些改变,在KYSE30和TE1细胞中过表达NORAD则效果相反(Fig 6C)。综上所述, NORAD/miR-224-3p/MTDH轴促进β-catenin的核积累,激活该蛋白,从而促进了ESCC细胞对CDDP的耐药和进展。
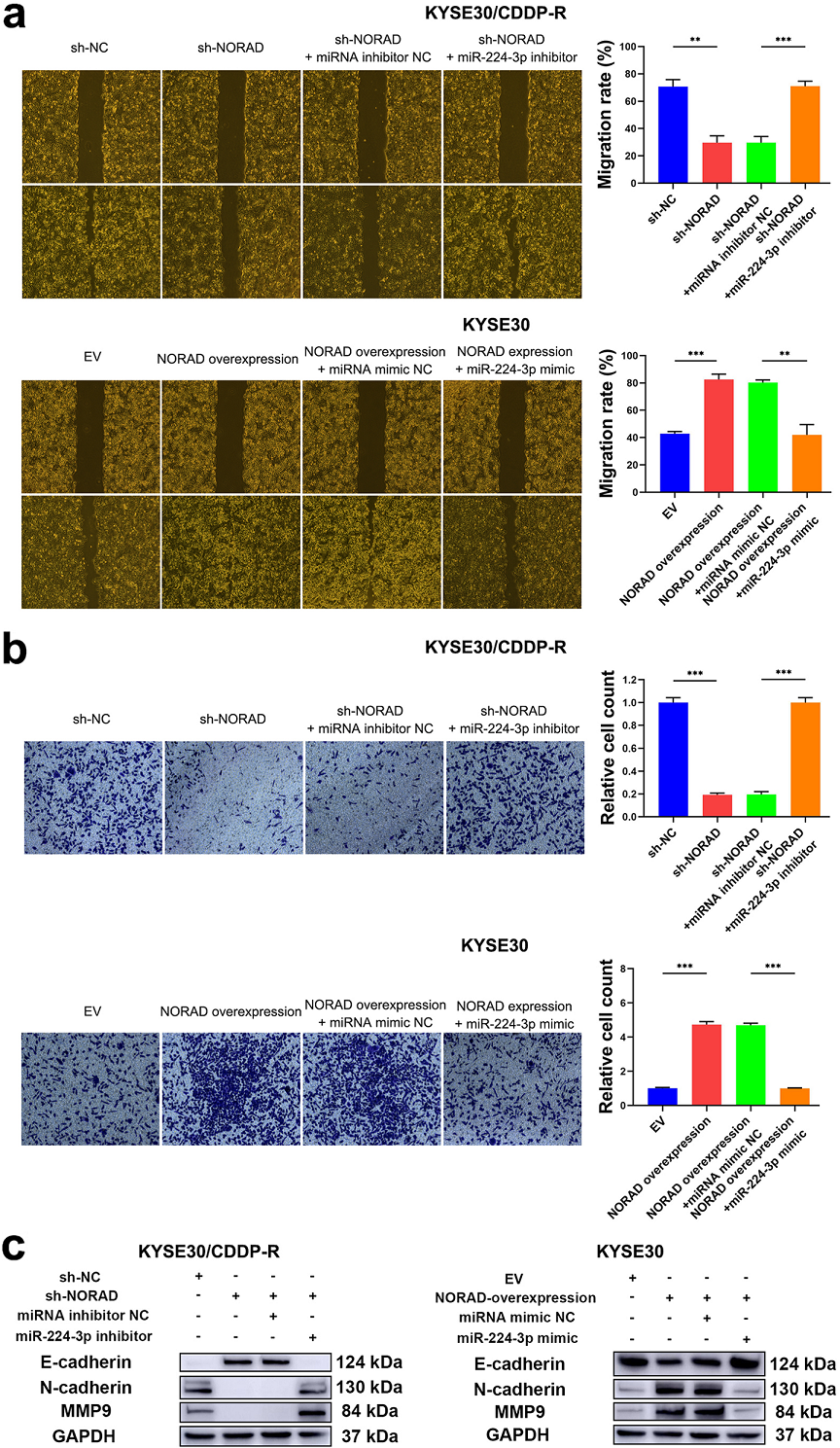
Fig 6 NORAD/miR-224-3p/MTDH轴促进ESCC细胞的进展
7. NORAD在体内促进CDDP的耐药和ESCC的生长
将8只雌性ESCC小鼠随机分成2组注射对CDDP耐药和敏感细胞(用于注射的CDDP耐药食管癌细胞之前sh-NORAD或sh-NC处理,CDDP敏感的细胞过表达NORAD),一周后,给小鼠腹腔注射指定浓度的CDDP或PBS。结果发现,耐药细胞来源的异种移植瘤,经CDDP处理后肿瘤体积明显小于对照组, MTDH表达降低,但肿瘤依然持续生长。第4周开始,转染NORAD敲低处理细胞的小鼠中注射CDDP的肿瘤显著消退,说明NORAD发挥了抑制作用(Fig 7a)。经NORAD过表达载体预处理的KYSE30细胞转染到小鼠中即使在CDDP处理下也能形成持续生长的肿瘤(Fig 7b)。接下来,作者通过免疫组化法检测sh-NORAD的KYSE30/CDDP-R细胞中的MTDH的表达量,MTDH的表达明显低于未处理的KYSE30/CDDP-R细胞。同样,在过表达NORAD 的KYSE30细胞的异种移植瘤中,MTDH的表达量高于EV细胞(Fig 7c)。在ESCC细胞中β-catenin的核表达与MTDH呈正相关(Fig 7d)。此外,CDDP诱导与γH2AX和cleaved caspase-3表达正相关,与异种移植肿瘤中MTDH呈负相关(Fig 7e-f)。 综上所述,这些结果表明,NORAD通过促进β-catenin的核积累,促进了ESCC细胞CDDP耐药性的进展和发展(Fig 7G)。
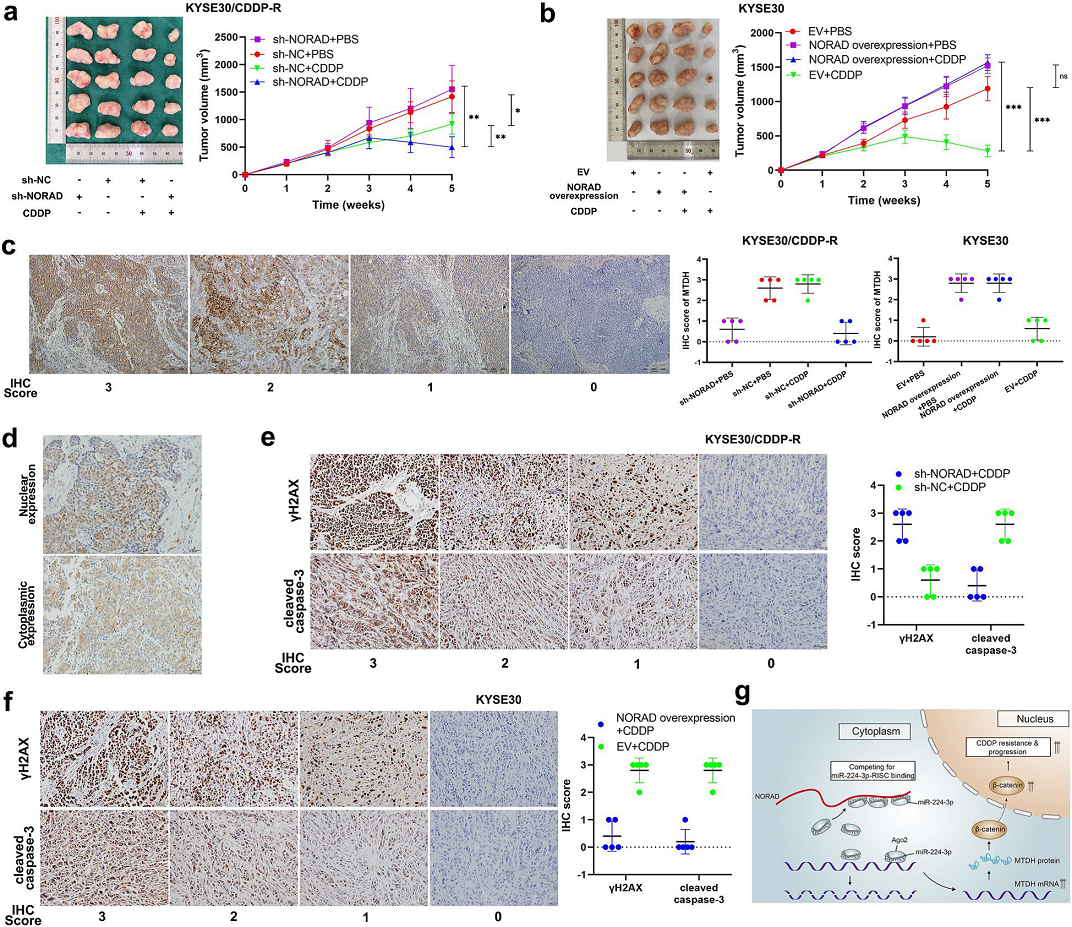
Fig 7 NORAD/miR-224-3p/MTDH轴有助于ESCC体内CDDP耐药
本研究中,作者发现NORAD是参与体外和体内ESCC CDDP耐药的关键lncRNA NORAD作为miR-224-3p海绵吸附miR-224-3p,降低miR-224-3p的表达水平,提高MTDH的表达量,促进β-catenin磷酸化,进而促进β-catenin的核积累,提高 ESCC细胞CDDP耐药性。
参考文献:
Jia Y.,et al.,Long non-coding RNA NORAD/miR-224-3p/MTDH axis contributes to CDDP resistance of esophageal squamous cell carcinoma by promoting nuclear accumulation of beta-catenin. Mol Cancer,2021. 20(1): p.162.