






LncRNA-HGBC通过调节miR-502-3p/SET/AKT通路促进胆囊癌发展
LncRNA在很多癌症的发生和发展中起着重要作用,但在胆囊癌(GBC)中,很多新的LncRNA如何对疾病进行调节需进一步研究。本文中,作者通过芯片筛选出胆囊癌(GBC)患者中胆囊恶性肿瘤与周围良性组织微阵列中差异表达的LncRNA和mRNA,并阐明其对GBC细胞增殖、迁移、侵袭的作用机制。
本文技术路线如下:

主要结果如下:
1 、鉴定出LncRNA-HGBC,并发现LncRNA-HGBC表达水平与GBC预后相关
为找出胆囊恶性肿瘤与周围良性组织中差异表达的LncRNA和mRNA,通过芯片技术筛选出一些候选LncRNA,并发现其中的LncRNA-HGBC(LncRNA Highly expressed in GBC)在GBC肿瘤细胞中表达量显著高于周围良性组织,且LncRNA-HGBC高表达的患者总生存率(OS)显著低于LncRNA-HGBC低表达的患者(Fig 1A and B),揭示了LncRNA-HGBC在胆囊癌肿瘤细胞转移中起积极作用。随后,作者通过RACE快速扩增得到LncRNA-HGBC的5′端及3′端序列(Fig 1C),再通过PCR 扩增GBC全长序列,Northern杂交进一步证实在GBC细胞系中LncRNA HGBC的RNA全长约为2 kb(Fig 1D)。为确定LncRNA-HGBC所在位置,作者分离了NOZ细胞的细胞核和细胞质部分,进行了qRT-PCR检测,初步判断LncRNA-HGBC主要定位在细胞质中,与原位杂交(FISH)结果一致(Fig 1E)。通过体外翻译实验,发现LncRNA-HGBC并未参与任何蛋白质的表达(Fig 1F),表明LncRNA-HGBC是一个不编码蛋白质的长链RNA。
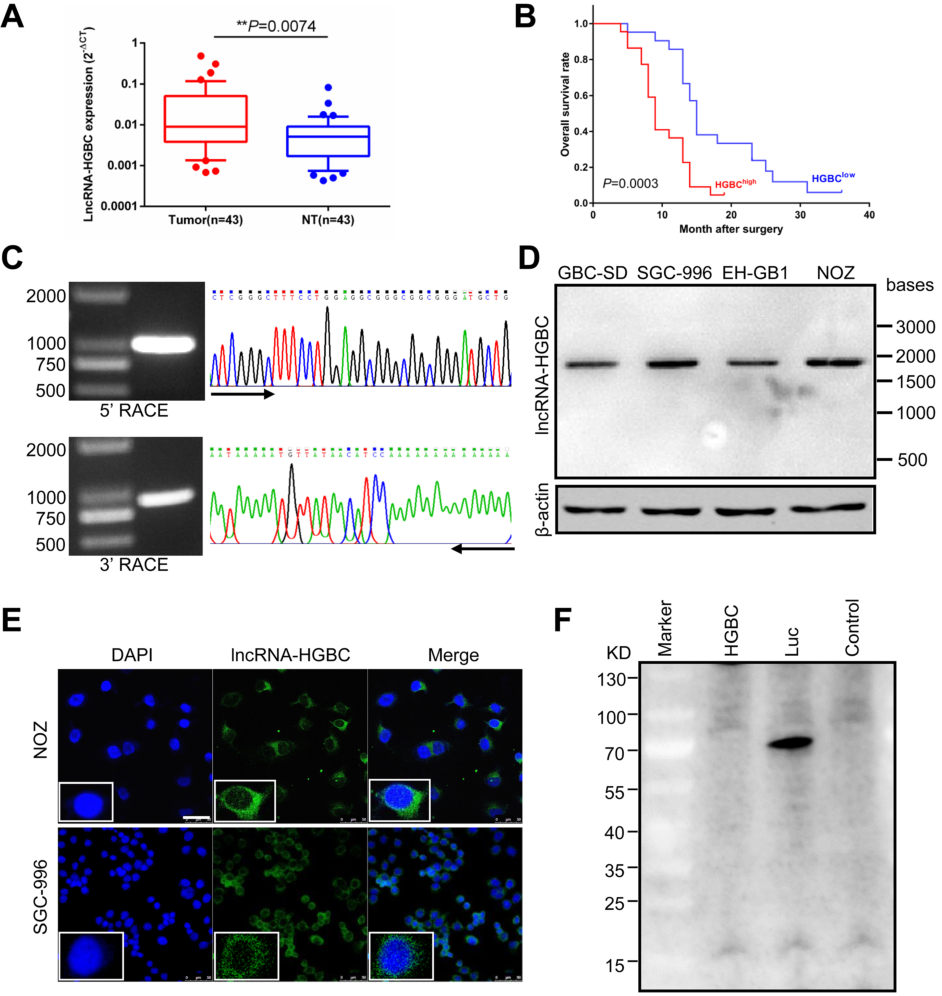
Figure 1 影响GBC进展的LncRNA的鉴定与表达分析
2、LncRNA-HGBC能在体内外促进GBC细胞增殖
为确定LncRNA-HGBC对GBC细胞增殖及对肿瘤发生的影响,研究中检测了LncRNA-HGBC在四种GBC细胞系(NOZ、GBC-SD、SGC-996、EH-GB1)中的表达情况,结果发现NOZ和SGC-996细胞系中LncRNA-HGBC的表达量高于GBC-SD和EH-GB1细胞系(Fig 2A)。在LncRNA-HGBC高表达的NOZ和SGC-996细胞系中通过干扰LncRNA-HGBC(Fig 2B),发现任意干扰两个区域都会使细胞增殖显著降低(Fig 2C),于此同时,干扰后菌落数量也显著减少,两种细胞系形成菌落的能力降低(Fig 2D)。此外,为了确定LncRNA-HGBC在体内对GBC细胞生长的影响,在小鼠中比较注射敲除了LncRNA HGBC的NOZ细胞和注射未经敲除的NOZ细胞时小鼠之间肿瘤差异,结果发现,前者较后者的肿瘤体积和肿瘤重量降低30%(Fig 2E)。通过免疫组化检测增殖细胞核抗原(PCNA),观察细胞增殖情况,也得到了相同的结论,与对照组比较,LncRNA hGBC水平降低会引起细胞增殖水平下降(Fig 2F)。另一方面,在GBC-SD 和 EH-GB1细胞系中,通过建立LncRNA hGBC过表达的发现过表达细胞系中细胞的增殖速度明显提高(Fig 2G 、2H、2I),此外,过表达细胞系中肿瘤的体积和重量是对照组的2.5倍(Fig 2J),通过IHC染色发现过表达细胞系异种移植瘤组织中增殖细胞核抗原(PCNA)上调(Fig 2K)。
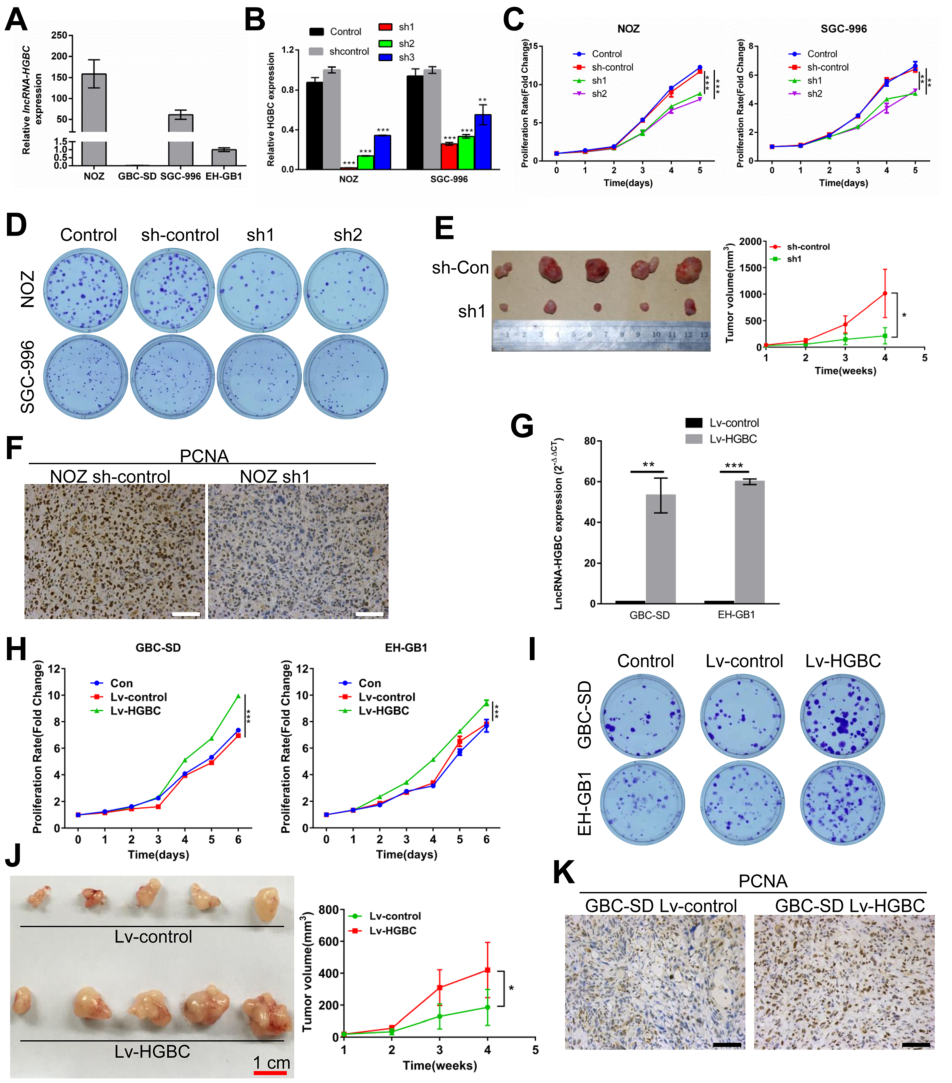
Figure 2 LncRNA-HGBC促进GBC细胞增殖和肿瘤生长
3、LncRNA -HGBC促进GBC细胞进行上皮细胞-间充质转化(EMT)和肿瘤转移
为了判断LncRNA –HGBC表达水平与淋巴结细胞迁移之间的关系,研究中首先提出了LncRNA –HGBC促进GBC细胞迁移的假设,并进行验证。通过transwell和基质胶侵袭实验,发现在NOZ 和 SGC-996 细胞系中,敲除LncRNA -HGBC可显著减少约30-40%的细胞迁移和侵袭(Fig 3A,3B)。反之,在GBC-SD 和 EH-GB1细胞系中细胞,对LncRNA- HGBC进行过表达,发现在GBC细胞迁移和侵袭能力增加20-50%(Fig 3C,3D)。EMT(上皮细胞-间充质转化)在很多肿瘤细胞迁移和侵袭中非常重要,为了评估EMT是否参与GBC侵袭,本研究中检测了两种EMT特异性标记物,分别是波形蛋白和N-钙粘蛋白。作者在NOZ 和SGC-996 cells细胞系中,敲除LncRNA –HGBC发现波形蛋白和N-钙粘蛋白的表达量降低(Fig 3E),反之,在GBC-SD 和 EH-GB1细胞系中,对LncRNA- HGBC进行过表达,发现波形蛋白和N-钙粘蛋白的表达量升高(Fig 3F),说明LncRNA- HGBC可能引起EMT的发生。为进一步验证,作者将NOZ细胞注射到小鼠脾脏中建立了裸鼠肝转移瘤模型,对照组的6只小鼠中有5只(5/6)在移植6周后显示荧光素酶信号增强,并在肝内发生转移,而注射 干扰LncRNA-HGBC后只小鼠中只有3只小鼠出现肝内转移。在肝脏中,对 LncRNA-HGBC 进行干扰的小鼠中转移灶较对照组降低10%(Fig 3G,3H)。
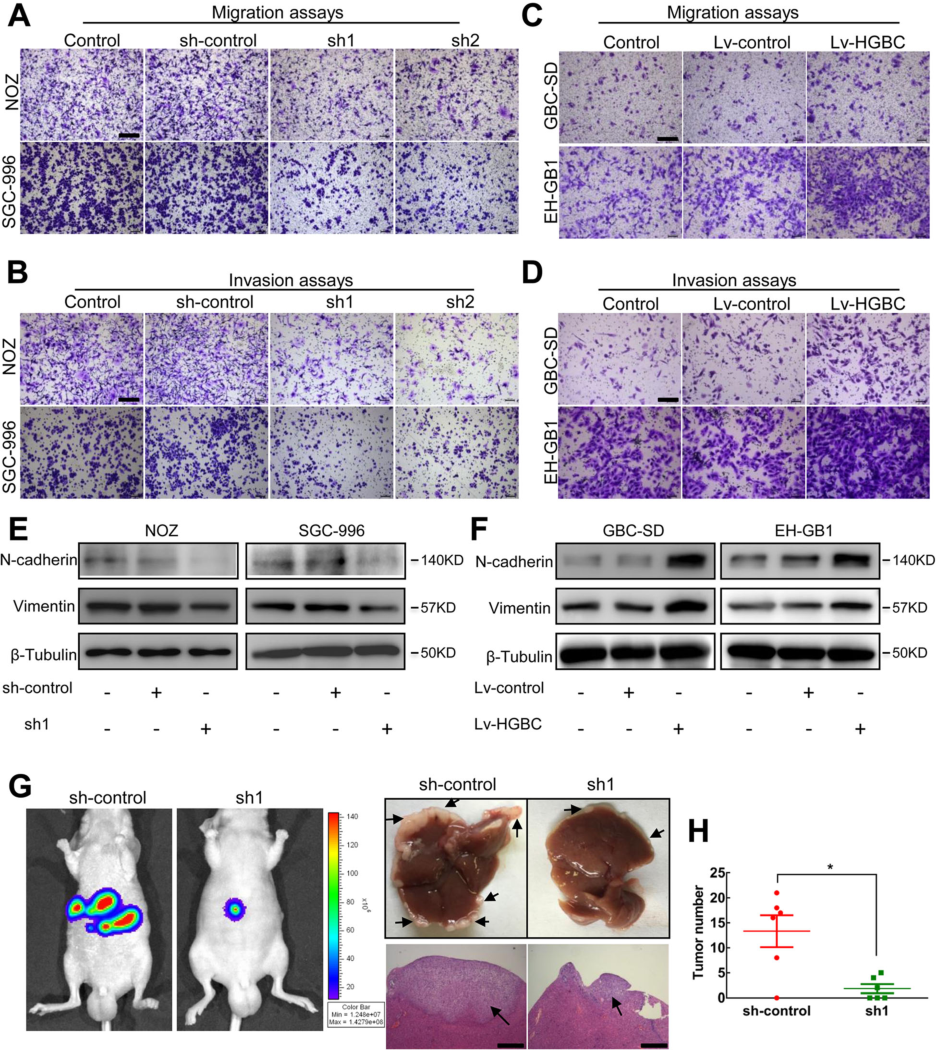
Figure 3 LncRNA-HGBC增强GBC细胞的侵袭能力
4、 HuR与LncRNA-HGBC特异结合并维持LncRNA-HGBC稳定表达
为探索LncRNA-HGBC与是否与GBC细胞存在潜在蛋白,通过pull-down实验,发现LncRNA-HGBC在37kd处存在与蛋白质的互作(Fig 4A),进一步通过 Western免疫印迹证实与LncRNA-HGBC结合的蛋白为HuR(Fig 4B),通过RNA结合蛋白免疫沉淀(RIP)验证LncRNA-HGBC与HuR的互作,与富含HuR结合位点的阳性对照EIF4E mRNA相比,通过RNA结合蛋白免疫沉淀(RIP)实验证验了LncRNA HGBC和HuR之间的特异性相互作用(Fig 4C)。为了进一步确定LncRNA HGBC中的哪个特定区域与HuR结合有关,作者构建了4个不同的LncRNA -HGBC缺失片段(Fig 4D),通过pull-down实验之后再进行Western免疫印迹确定了LncRNA -HGBC的1759–1906nt区域与HuR特异结合(Fig 4E)。为了阐明HuR是否影响LncRNA-HGBC在GBC细胞中的稳定性,通过两次HuR敲除实验均发现LncRNA-HGBC在NOZ细胞系中的表达量降低(Fig 4F);为进一步探索 LncRNA-HGBC表达量的降低是不是由于 LncRNA-HGBC衰退所致,通过阻断RNA的转录,观察30h内 LncRNA-HGBC表达量的变化,结果发现,随着HuR的逐步消耗,LncRNA-HGBC水平的半衰期从18小时降低到5小时(Fig 4G),这一系列实验表明HuR与LncRNA HGBC相互作用并维持LncRNA HGBC稳定表达。
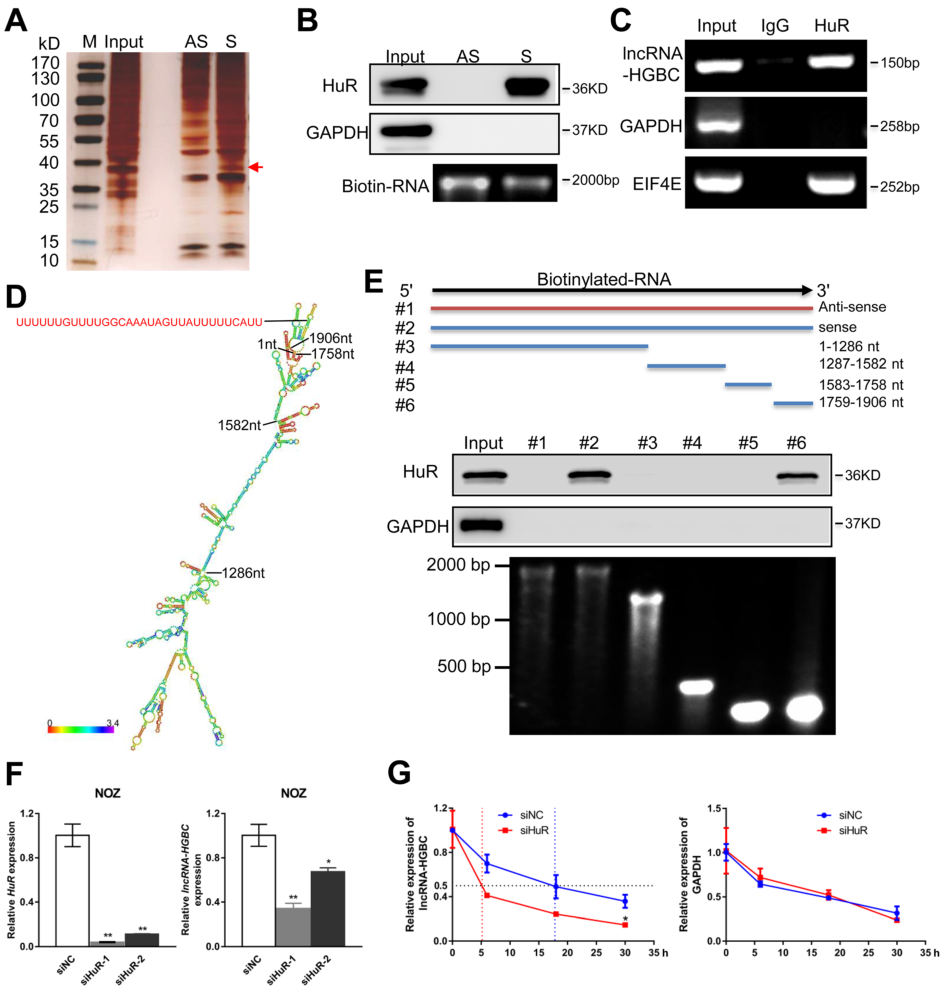
Figure 4 HuR与LncRNA-HGBC特异结合并有助于维持LncRNA-HGBC稳定表达
5、 LncRNA-HGBC作为一种竞争性内源性RNA抑制miR-502-3p表达
前期的研究已经证实LncRNA-HGBC定位在细胞质上,作者初步假设LncRNA -HGBC可能作为miRNA海绵影响miRNA的表达,在GBC的进展中发挥作用。为了找出与LncRNA-miRNA相互作用的miRNA, 作者将LncRNA-HGBC全长序列克隆到pmirGLO双荧光素酶报告载体中,将6个与肿瘤抑制相关的候选miRNA通过双荧光素酶实验进行分析,发现其中的miR-502-3p和miR-618可抑制LncRNA-HGBC的荧光素酶活性(Fig 5A),说明LncRNA-HGBC可能与miR-502-3p、miR-618相互作用。作者根据先前的miRNA微阵列结果发现,与邻近的非肿瘤组织相比,在GBC组织中miR-502-3p表达量确实下调了。当LncRNA-HGBC中1176 - 1183nt之间的序列被敲除后将不能与miR-502-3p结合,荧光活性将不会再被抑制(Fig 5B)。AGO2蛋白可以在转录后调控调节miRNA丰度,内源性Ago2的减少会降低成熟miRNA的表达和活性。进一步进行免疫共沉淀实验,发现AGO2抗体复合物在LncRNA-HGBC和miR-502-3p中特异富集,表明miR-502-3p是一个真正的靶向LncRNA-HGBC的miRNA(Fig 5C)。为进一步确定LncRNA-HGBC是否与miR-502-3p结合,作者通过MS2-RIP找到与LncRNA-HGBC结合的 miRNA,结果再一次证实了miR-502-3p与LncRNA-HGBC特异性结合(Fig 5D),而且miR-502-3p结合位点的突变也使miR-502-3p与LncRNA-HGBC之间的结合不再成立,也验证了LncRNA-HGBC和miR-502-3p之间的直接结合。为了进一步阐明LncRNA-HGBC和miR-502-3p之间基因表达的调控关系,评估了不同lncRNA-HGBC表达水平对细胞中的miR-502-3p水平的影响。结果发现,在 NOZ细胞系中,敲低LncRNA-HGBC 会导致miR-502-3p表达量上调了60%(Fig 5E左图),在EH-GB1细胞系中,LncRNA-HGBC过表达后miR-502-3p的表达被抑制,表达量下降了50%(Fig 5E右图), 但对miR-502-3p进行敲除或过表达时LncRNA-HGBC的表达量没有变化,说明LncRNA-HGBC作为海绵会抑制miR-502-3p表达,但不能诱导其降解。为了检测LncRNA HGBC的活性是否取决于其与miR-502-3p的结合,在异位表达野生型LncRNA HGBC和结合突变型HGBC-MUT后进行cck-8和Transwell分析,结果发现,LncRNA-HGBC(而非HGBC-MT)的表达可显著促进细胞迁移、侵袭和增殖(Fig 5F-G),由此可见,LncRNA-HGBC与miR-502-3p能结合,并可作miR-502-3p的海绵抑制其表达。
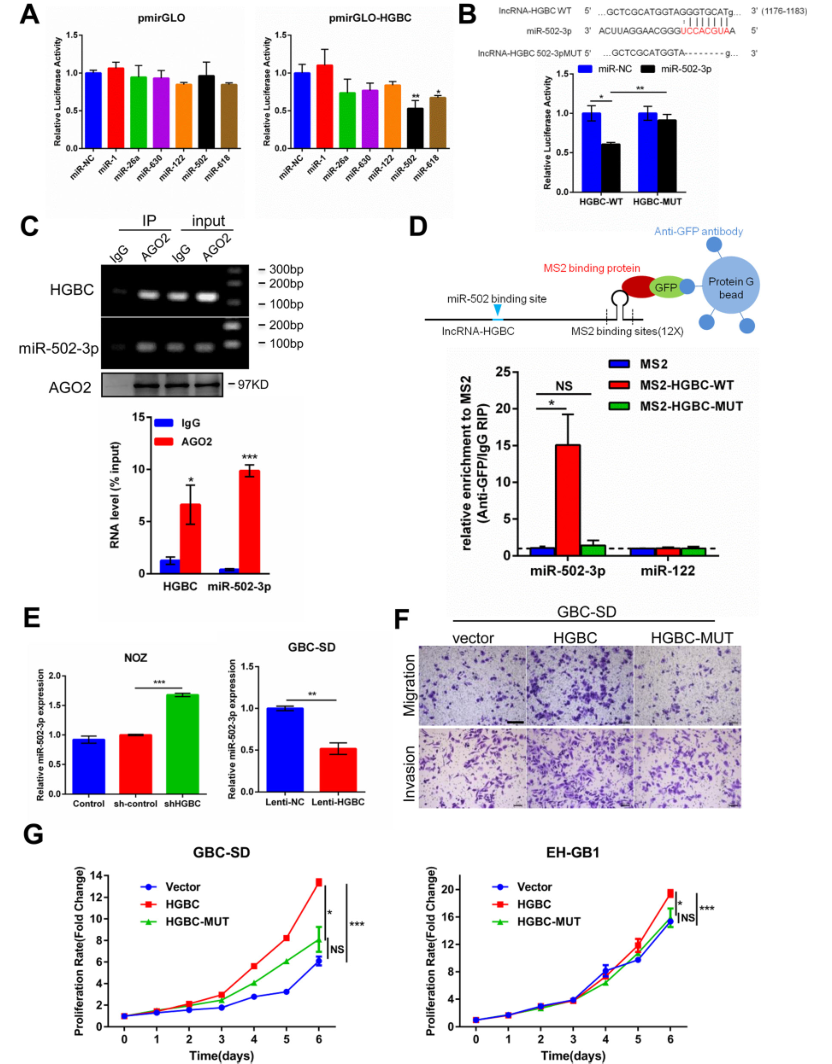
Figure 5 LncRNA-HGBC作为一种竞争性内源性RNA能直接与miR-502-3p结合
6、LncRNA-HGBC通过竞争性结合miR-502-3p调节SET的表达
miR-502-3p可以靶向多种蛋白质,并在癌症发展过程中发挥重要,作者通过芯片数据分析发现SET在GBC组织中也上调,那么miR-502-3p是否对SET进行调节需进一步探索。作者将含有SET 3′UTR的荧光素酶报告载体转染到293T细胞中,然后在转染miR-502-3p模拟物时评估荧光素酶活性,发现miR-502-3p显著降低了SET报告载体的荧光素酶活性此外,在GBC细胞系中,对miR-502-3p进行过表达或敲除之后,SET随之变化,说明SET是miR-502-3p的直接靶点。
由于LncRNA-HGBC和SET 3′UTR都与miR-502-3p结合,于是进一步研究LncRNA HGBC是否调节了GBC细胞中miR-502-3p,并对SET进行抑制。将pmirGLO-SET荧光素酶报告载体与LncRNA-HGBC过表达质粒进行共转染,过表达miR-502-3p将会抑制SET报告载体的荧光素酶活性,但过表达cRNA-HGBC将会诱导SET报告载体的荧光素酶活性(Fig 6A)。当miR-502-3p和LncRNA-HGBC同时被添加时,荧光活性恢复到对照水平,这就意味着LncRNA- HGBC能够通过与miR-502-3p结合,进而上调SET。进一步研究SET 如何控制蛋白表达,在内源性LncRNA HGBC 高表达的 NOZ细胞系中进行LncRNA-HGBC干扰和miR-502-3p抑制,结果发现LncRNA-HGBC的敲除抑制了SET的表达,miR-502-3p抑制剂的加入反而促进了SET的表达(Fig 6B,左图)。然而,在内源性LncRNA HGBC 低表达的EH-GB1细胞系中,过表达LncRNA-HGBC诱导SET表达,此时加入miR-502-3p,SET表达量将会降低至对照水平(6B,右图)。在NOZ细胞细胞系中,对LncRNA HGBC进行干扰将会抑制细胞增殖、迁移和侵袭能力,而miR-502-3p的加入逆转了这种局势(Fig 6CandD) 。
然而,在EHGB1细胞系中,过表达miR-502-3p延缓了细胞的生长、迁移和侵袭(Fig 6C and E)。为检测LncRNA HGBC是否诱导SET表达是否取决于LncRNA HGBC与miR-502-3p的结合,在GBC-SD和EH-GB1两个细胞系中,通过过表达LncRNA-HGBC发现mRNA和蛋白质水平上SET表达上调,LncRNA HGBC的过度表达导致SET表达增加,而LncRNA HGBC的miR-502-3p结合位点突变却未能诱导SET表达(Fig 6FandG),通过IHC发现,肿瘤异种移植模型中在LncRNA-HGBC耗尽之后SET的表达量也随之降低,过表达LncRNA-HGBC的肿瘤异种移植模型中LncRNA-HGBC表达量显著增加,说明LncRNA-HGBC和SET之间存在共表达,LncRNA-HGBC通过竞争性结合miR-502-3p调节SET的表达。
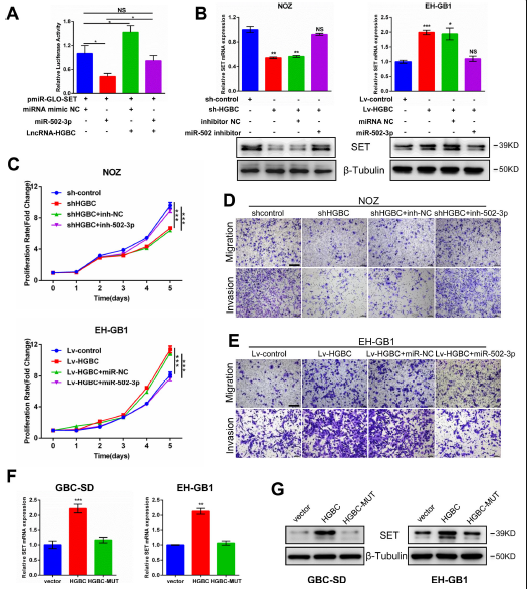
Figure 6 LncRNA-HGBC通过竞争性结合miR-502-3p调节SET的表达
7、AKT是SET的下游效应物、lncRNA与miR-502-3p、SET和p-AKT的关系
接下来对LncRNA-HGBC和SET对影响GBC转移的分子机制进行研究。研究发现对SET进行敲除后GBC细胞的增殖和侵袭能力明显降低,已有研究表明SET在很多癌症中是通过激活AKT提高癌细胞活性,那么LncRNA-HGBC-miR-502-3p-SET-AKT是否作为一个完整的信号通路介导GBC发生和转移的需要进一步验证。
作者首先检测了AKT激酶的活性,在NOZ和SGC-996细胞系中,通过WB实验发现了敲除LncRNA -HGBC显著抑制了AKT的活性(Fig 7A);在GBC-SD和EH-GB1细胞系中,过表达LncRNA-HGBC能诱导 AKT的磷酸化,起到信号传导的作用(Fig 7B)。
为确定LncRNA-HGBC对AKT的诱导是否通过调控miR-502-3p实现的,在GSC-SD和EH-GB1细胞系中,作者通过突变LncRNA-HGBC上与 miR-502-3p结合的位点,发现过表达LncRNA-HGBC能诱导 AKT的磷酸化,以及N-钙粘蛋白、波形蛋白的表达,进行突变的LncRNA-HGBC不能再对蛋白进行诱导(Fig 7C)。在EH-GB1细胞系中,将miR-502-3p导入过表达LncRNA-HGBC中会降低了这些蛋白的表达(Fig 7D);MK2206作为AKT抑制剂能逆转LncRNA-HGBC对N-钙粘蛋白和波形蛋白表达的影响(Fig 7E),表明LncRNA HGBC需要AKT激活。
为了进一步分析LncRNA-HGBC-miR-502-3p-SET-AKT之间的关联,作者在43对GBC组织中进行分析,发现LncRNA HGBC与miR-502-3p呈负相关,与SET和HuR mRNA水平呈正相关(Fig 7F),与非肿瘤组织相比,HuR在GBC组织中表达上调,通过IHC分析发现,与非肿瘤组织相比,SET或p-AKT在GBC组织中表达上调(Fig 7G),此外,LncRNA -HGBC表达与SET和p-AKT表达呈正相关(Fig 7H and G)。综上所述,LncRNA HGBC能通过竞争性结合miR-502-3p发挥作用,然后激活下游SET和AKT表达,从而加强肿瘤细胞的侵略性。
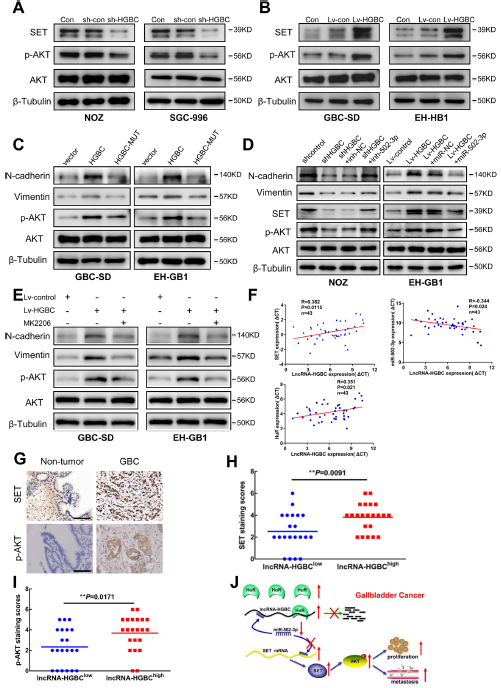
Figure 7 AKT是SET的下游效应物、lncRNA与miR-502-3p、SET和p-AKT的关系
本文发现了一个与GBC预后差相关的LncRNA-HGBC,并阐明了LncRNA-HGBC-miR-502-3p-SET-AKT通路介导GBC增殖、迁移、侵袭的分子机制。
参考文献:
Hu Y P , Jin Y P , Wu X S , et al. LncRNA-HGBC stabilized by HuR promotes gallbladder cancer progression by regulating miR-502-3p/SET/AKT axis[J]. Molecular Cancer, 2019, 18(1). DOI:10.1186/s12943-019-1097-9.